細菌細胞結構
圖1:細菌細胞結構
細菌屬于真核細胞,有核孔復合物,胞漿中有細胞骨架微絲及各類細胞器
1.細胞壁
細胞壁主要物質為碳水化合物,包括一些晶狀物質,如α-葡聚糖、幾丁質、β-(1,3)葡聚糖聯合的N-甲基糖胺聚合物。其它如甘露聚糖等。
(幾丁質:細菌螢光染色的主要成份;甘露聚糖-GM實驗主要用于曲霉的血漿學測量)
2.細胞膜
包括磷脂、麥角膽固醇、葡聚糖合成酶等
主要系統性抗細菌抗生素
1.麥角膽固醇的合成
唑類和多烯類抗細菌抗生素通過影響麥角膽固醇起到抗細菌作用。(麥角膽固醇的合隔閡圖2)
圖2:麥角膽固醇的合成及抗細菌抗生素作用
2.唑類作用機制
麥角膽固醇是好多細菌的細胞膜內酯的主要成份。通過抑制14α-血紅素去酰基酶(羊絨內酯去酰基酶),這是一種細菌細胞色素P450(P450,CYP)依賴酶,唑類抗生素使細胞膜的麥角膽固醇不能正常合成,損傷細菌細胞膜,造成細菌死亡。
圖3唑類抗生素物理式
14α-去酰基酶位點與唑類抗生素的結構主要影響了抗生素與靶酶的親和性,對一些細菌種類,唑類可形成交叉耐藥。伊曲康唑和泊沙康唑延長的非極性基團加大了與14α-去酰基酶的結合,所以抑菌譜也更寬。伏立康唑是氟康唑的衍生物,但其結構上有一個α-0-group,所以可以抗曲霉和其他球狀細菌。
三唑類抗生素耐藥主要因為14α-去酰基酶上與唑類結合位點的改變(azoleof14α-),氟康唑耐藥與MDR1外排泵的過抒發外有關,外排泵CDR1及CDR2可使唑類交叉耐藥。克柔念珠菌對氟康唑的天然耐藥與抗生素和14α-去酰基酶的結合破損有關細胞膜選擇性通透,而新的唑類強化了這些結合。光滑念珠菌對氟康唑耐藥一般是外排泵過抒發的結果,因而,常常形成唑類的交叉耐藥。
3.特比萘芬作用機制:
與唑類相像,特比萘芬抑制角鯊烯環氧化酶來抑制麥角膽固醇的合成,特比萘芬主要分布在皮膚和甲床中細胞膜選擇性通透,而血藥含量相對較低,通常用于甲癬和皮膚細菌病。
4.兩性霉素B作用機制:
AMB直接和麥角香豆素結合產生復合體,并產生孔道,造成細胞內物質泄露。兩性霉素B對麥角膽固醇豐富的細胞膜以及甘油三酯豐富的細胞膜(喂奶植物及人)均有影響,所以可導致毒副作用,例如在心臟富集,導致腎損傷。兩性霉素B也可以使促炎細胞因子釋放,致使輸注過程的發熱,寒顫。兩性霉素B含脂復合劑型可降低心臟分布,毒副作用減少。AMB臨床耐藥罕見,固乙醇的取代以及形成中和酶來耐受氧損傷是AMB天然耐藥及獲得性耐藥的主要機制。
5.棘白菌素作用機制:
棘白菌素作用于細菌細胞壁,通過抑制β-(1,3)-d-葡聚糖合成酶來抑制細胞壁主要結構之一的β-(1,3)-d-葡聚糖的合成發揮作用。細胞壁中β-1,3-d-葡聚糖合成過程及酶的抒發主要決定了棘白菌素的抗細菌譜。棘白菌素對念珠菌屬及曲霉菌屬有較好活性。耐藥罕見,「熱點」地帶FKS1及FKS2突變造成的β-(1,3)-d-葡聚糖合成酶的主要催化亞基改變可引起棘白菌素作用增加,MICs下降以及醫治失敗。
6.氟胞吡啶:
氟胞噻吩經胞吡啶透酶步入細菌細胞,經胞吡啶脫氨酶代謝為氟尿嘧啶,取代尿吡啶造成細菌RNA的錯誤編碼。但是,肝臟菌群也可以把5-氟胞吡啶轉變為5-氟尿嘧啶,致使難受、嘔吐、腹瀉以及骨髓抑制等不良反應。而胞吡啶透酶、胞吡啶脫氨酶突變容易發生,所以氟胞吡啶通常僅用于聯合診治。
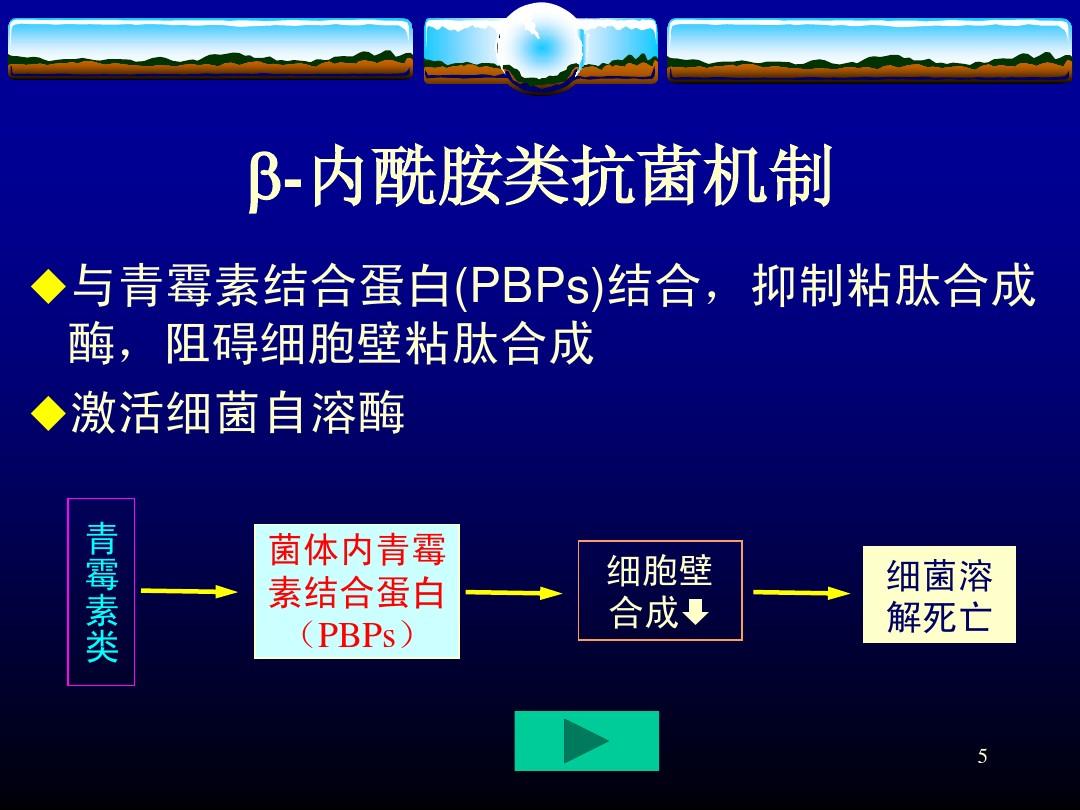
圖4.系統性抗細菌抗生素作用機制示意圖
抗生素的抗細菌活性比較
抗細菌抗生素的不良反應
參考書籍:
1.《》
2.《實用抗感染醫治學》
3.《醫學原蟲學-實驗室檢驗手冊》
參考文獻:
1.LewisRE.in.[J].MayoMayo,2011,86(8):805-17.
2.XieJL,PolviEJ,T,etal.druginhuman.[J].,2014,9(4):523.
3.DW,MJ.Howtothe[J].,2015,347(6229):1414-1416.
4.A,R,CampaM,etal.basisoftoazole.[J].in,2002,8(2):76-81.