好醫師編者按
細胞膜( cell )也稱質膜( ) ,是分隔細胞質與細胞周圍環境的一層膜結構,厚度7~8nm。細胞膜和細胞內各類細胞器的膜結構及其物理組成是基本相同的,主要由脂類和蛋白質組成,還有少量脂類物質。
其中,和脂類的比列在不同種類的細胞可相差很大。一般而言,在功能活躍的細胞,膜蛋白濃度較高;而在功能簡單的細胞,膜蛋白濃度相對較低。例如,膜蛋白與膜脂類在小腸粘膜上皮細胞膜中的重量比可高達4.6:1,而在構成神經纖維髓鞘的施萬細胞膜中的重量比僅為0.25:1。關于各類化學成分在膜中排列的方式,目前廣為接受的是和于1972年提出的液態鑲嵌模型(fluid model)學說。這一學說覺得,液態脂類單層構成膜的基架,不同結構和功能的蛋白質鑲嵌在其中,糖類分子與脂類、蛋白結合后附在膜的外表面(圖2-1)。
(一)細胞膜的脂類
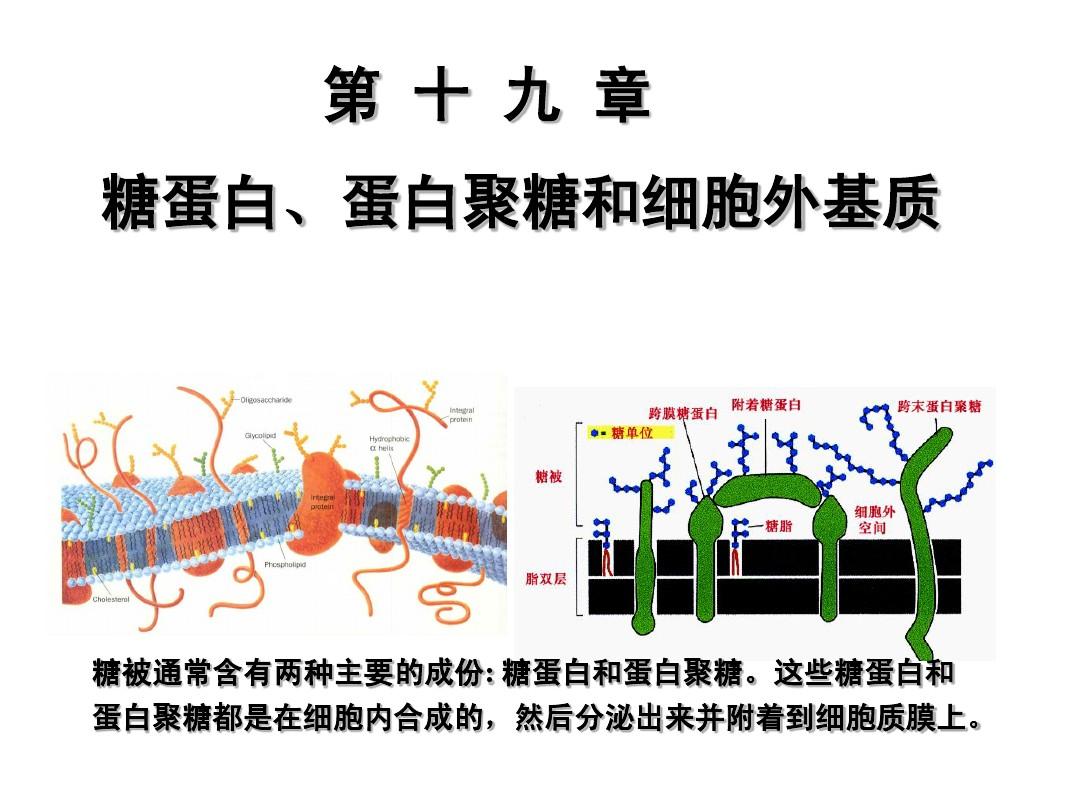
在多數細胞,雖然膜蛋白質總重量小于脂類,但因為蛋白質的分子量比脂類大得多,所以脂類的分子數卻遠少于蛋白質。因而,脂質成為細胞膜的基本架構,連續包被在整個細胞的表面。膜脂類主要由磷脂( )、和少量糖脂( )構成。在大多數細胞的膜脂類中,磷脂占總數的70%以上,膽固醇不超過30% ,糖脂不超過10%。磷脂是一類富含乙酸的單糖。磷脂中濃度最多的是磷脂酰膽堿,其次是磷脂酰絲氨酸和磷脂酰乙醇胺,含量最低的是磷脂酰肌醇。各種膜脂類在膜上的分布是不對稱的。大部分的磷脂酰膽堿和全部的糖脂都分布在膜的內層,含多肽的磷脂(磷脂酰絲氨酸、磷脂酰乙醇胺、磷脂酰肌醇)主要分布在膜的外層。其中,磷脂酰肌醇的濃度盡管甚少,但可作為細胞內第二信使三磷酸肌醇( IP,)和二酰甘油( DG)的供體,因而在跨膜信號轉導中有重要作用(見第二節)。
脂質分子都是雙嗜性分子( )。例如,磷脂分子中富含乙酸和核苷酸的頭端具有親水性,含有較長脂肪酸的尾端具有疏水性;膽固醇分子中的甲基以及糖脂分子中的糖鏈具有親水性,分子的另一端則具有疏水性。脂質分子的雙嗜特點使之在質膜中以脂類單層()的方式存在,即兩層脂類分子的親水端分別朝向細胞外液或胞質,疏水的脂肪酸烴鏈則彼此相對,形成膜內部的疏水區。質膜的疏水區是水以及水溶性物質如葡萄糖和各類帶電離子的天然屏障,但脂溶性物質如氫氣、以及丙酮等則很容易穿透。脂質分子呈單層排列是和于1925年通過對紅細胞膜化學成分的測定和紅細胞膜表面積的估算猜想下來的。
膜脂類可因體溫改變而呈凝膠或溶膠狀態。正常人溫度低于膜脂類的熔點,即已超過兩種狀態的轉換氣溫,故膜脂類在人體內呈溶膠狀態,具有一定程度的流動性。脂質雙分子層在熱力學上的穩定性和它的流動性,使細胞還能承受相當大的張力和變型而不至于斷裂,如紅細胞有很強的變形性和可塑性,能通過比其半徑還小的毛細血管和血竇縫隙。膜脂類的流動性還可使嵌入的膜蛋白發生側向聯通、聚集和相互作用。細胞的許多基本活動,如膜蛋白的相互作用、膜泡運輸、細胞的運動、分裂、細胞間聯接的產生等都有賴于質膜保持適當的流動性。除了與氣溫有關外,質膜的流動性還與膜脂類的成份及膜蛋白的濃度有關。膽固醇分子具有不易變型的甾環結構,后者與脂肪酸鏈的結合可限制脂類的流動,故膜脂類中尿酸濃度愈高,膜的流動性就愈低;脂肪酸烴鏈寬度愈長、飽和脂肪酸愈多,膜的流動性也愈低,如植物脂肪以飽和脂肪酸為主,室溫下可呈固態;膜中鑲嵌的蛋白質濃度增多時也會增加膜的流動性。
(二)細胞膜的蛋白
細胞膜的功能主要是通過膜蛋白( )實現的。根據膜蛋白在膜上的存在形式,可將其分為表原液蛋白( ) 和整合膜蛋白( ) 兩類(見圖2-1)。
表原液蛋白約占膜蛋白總數的20%~30%,主要附著于細胞膜的內表面。例如,膜骨架蛋白和錨定蛋白。前者屬于結構蛋白,常以蛋白分子多聚體產生的長條細絲方式出現,可使質膜具有一定的硬度和彈性;后者用于整合膜蛋白的定位,將其固定到特定的質膜位置。有的表面蛋白還以酶的方式發揮胞內訊號轉導作用或參與個別物質轉運的控制。表面蛋白通過靜電引力與脂類的親水部份相結合或通過離子鍵與膜中的整合蛋白相結合,但其結合力較弱。高鹽堿液可使離子鍵斷掉,因而可用于表面蛋白從膜中的洗脫。
整合膜蛋白約占膜蛋白總數的70%~80%,它們以其肽鏈一次或反復多次穿越膜脂類單層為特點。穿越脂類單層的肽段以疏水性氨基酸殘基為主,肽鍵之間易產生構象,因而多以α螺旋結構的方式存在;露出膜外表面或內表面的肽段則是親水性的細胞膜結構細胞膜結構,分別以直鏈方式構成聯接疏水性α跨膜螺旋的細胞外環或細胞二環。一個α跨膜螺旋需18~21個氨基酸殘基能夠穿越厚約3nm的疏水區。因此,可依照肽鏈中所含的有足夠寬度的疏水性片斷的數量來推斷蛋白是否為跨膜蛋白及其跨膜次數。例如,G蛋白耦聯受體的肽鏈包含7個有足夠寬度的疏水性片斷,因而猜測它是一個7次跨膜的蛋白。整合膜蛋白與脂類分子結合緊密,在膜蛋白純化過程中可用兩性洗滌劑使之與脂類分離。一般來說,與物質跨膜轉運功能和受體功能有關的蛋白都屬于整合膜蛋白,如載體、通道、離子泵、G蛋白耦聯受體等。也有一些整合膜蛋白作為黏附分子()在細胞與基質、細胞與細胞之間發揮作用。
各種功能蛋白質分子在質膜中的位置分布存在區域特點,這與細胞完成其特殊功能有關。例如,骨骼肌細胞膜上的N2型乙酰膽堿受體一般都集中在與神經末梢相對應的終板膜上,這與神經胸肌之間的信息傳遞有關;有髓神經纖維軸突膜上的電流門控鈉通道幾乎全部集中在裸露的郎飛結處,這與激動的發生和跳躍式傳導(見后)有關。
(三)細胞膜的脂類
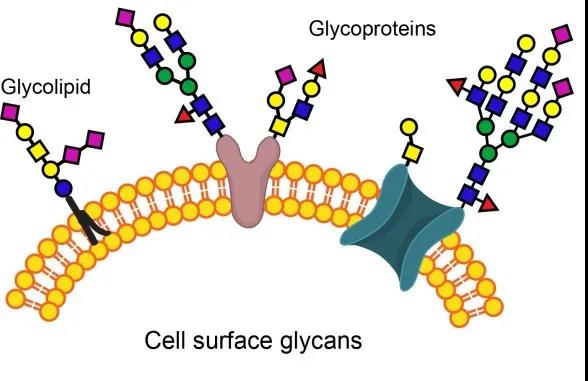
細胞膜中的脂類主要是一些甾醇和多糖鏈,它們以共價鍵的方式與膜蛋白或膜脂類結合而產生糖蛋白()或糖脂( )。大多數整合蛋白都是糖蛋白,近1/10的膜脂類是糖脂。結合于糖蛋白或糖脂分子上的糖鏈幾乎總是伸向細胞膜的內側(被稱為細胞“天線”),形成細胞的糖包被()。細胞表面糖包被的互相接觸可促使細胞之間的互相接觸和作用;許多脂類帶有負電荷,這促使細胞表面呈現負電性,從而抵觸帶有負電荷的物質與其接觸,如血液中紅細胞之間才能保持分開狀態就與膜上唾液酸(一種天然的碳水化合物)攜帶著負電荷有關;許多脂類還作為一種分子標記發揮受體或抗體的功能。例如,霍亂毒素的受體就是一種稱為Gm1的糖脂;紅細胞膜上ABO血型系統的抗體就是由糖蛋白或糖脂上不同的寡糖鏈所決定的,其中b型抗體和a型抗體的差異僅在于此糖鏈中一個糖基的不同(見第三章)。