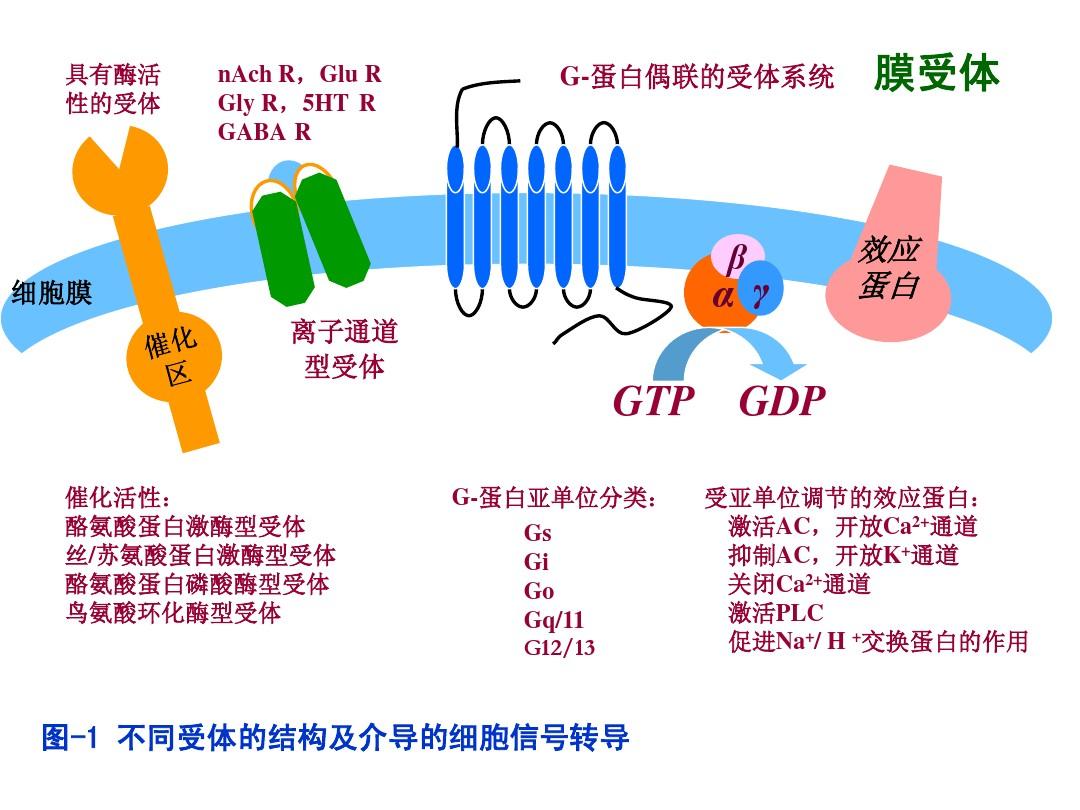
1.膜表面免疫球蛋白(mIg)這是B細胞特異性辨識抗體的受體,也是B細胞重要的特點性標志。不成熟B細胞抒發mIgM,成熟B細胞又抒發了mIgD,即同時抒發mIgM和mIgD,有的成熟B細胞表面還mIgG、mIgA或mIgE。在B細胞分化過程中細胞膜受體,前B細胞的胞漿中可有IgM的重鏈μ鏈,但無mIgM;當發育為不成熟B細胞時,胞漿中μ鏈消失,胞膜上開始抒發mIgM。在單個B細胞表面所有Ig的可變區都由相同的VH和VL基因所編碼,因而它們的奇特型和結合抗體的特異性是相同的。抗體剌激后的B細胞mIgD很快消失,記憶B細胞表面不存在mIgD。作為B細胞受體(BcellBCR)的mIgM外,還有Igα和Igβ兩種氨基酸鏈,分別命名為CD7979a和CD7979b,共同與mIg產生BCR復合物。
2.補體受體(CR)
B細胞膜表面具有CR1和CD2。CR1(CD35)可與補體C3b和C4b結合,進而促使B細胞的活化。CD2(CD21)的官能團是C3d,C3d與B細胞表面CR2結合亦可調節B細胞的生長和分化。
3.EB病毒受體
CR2(CD21)也是EB病毒受體,這與EB病毒選擇性感染B細胞有關。在體外可用EB病毒感染B細胞,可使B細胞永生化()而建成B細胞母細胞樣細胞株,在人單克隆抗原技術和免疫學中有重要應用價值。在體內,EB病毒感染與傳染性單核細胞增多癥、氏淋巴瘤以及癌癥等的發病有關。
4.致有絲分裂原受體歐洲桑寄生絲分裂原(pokeweedPWM)對T細胞和B細胞均有致有絲分裂作用。在大鼠,脂寡糖(LPS)是常用的致有絲分裂原。據悉金紅色獼猴桃球菌種(SAC)因富含金紅色獼猴桃桿菌A蛋白(ASPA),可通過與mIg結合剌激人B細胞的增殖。據悉細胞膜受體,小麥凝集素(SBA)可凝集B細胞。
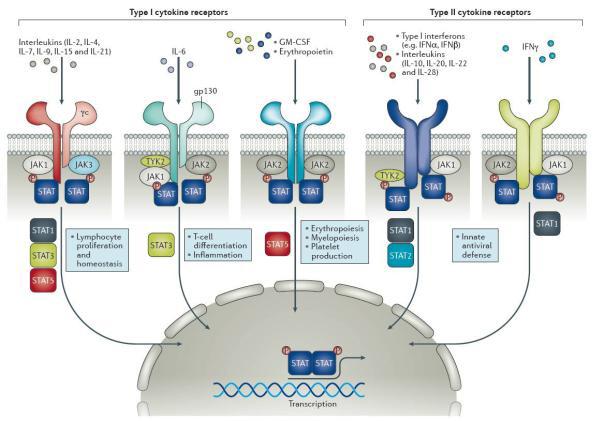
5.細胞因子受體
多種細胞因子調節B細胞的活化、增殖和分化是通過與B細胞表面相應的細胞因子受體結合而發揮調節作用的。B細胞的細胞因子受體主要有IL-1R、IL-2R、IL-4R、IL-5R、IL-6R、IL-7R、IL-11R、IL-12R、IL-13R、IL-14R、IL-γR、IL-αR和TGF-βR等。