動物的味道受體蛋白在細胞膜上產生離子通道,受體與異味分子結合時通道打開。解析非激活和激活狀態下通道的結構有助于深入了解動物怎樣測量和分辨不同的味道。
當使用驅蟲劑來制止動物傳播的病癥時,我們可能會思索這種物質是怎樣起作用的,它們如何能夠更有效。避蚊胺是驅蟲劑中最常用的化合物,被覺得可以廣泛激活動物的味道受體蛋白,攪亂動物找尋寄主的味覺編碼[1]。但尚不清楚避蚊胺或天然味道分子怎樣結合并影響動物味道受體的活性。在《自然》發表的一篇文章中,delMármol等人[2]報導了與避蚊胺或丁香酚相關的動物味道受體的結構,為味道怎樣與受體結合,以及激活受體的結構隨即怎樣變化提供了關鍵看法。
不同植物的味覺系統早已進化到可以完成高度專業化的任務。果蠅專精霉爛的香蕉,蚊蟲找尋寄主,而人類可以分辨大量與食物有關的異味。腰部植物和無肋骨植物都使用一組諸多味道受體來測量和分辨數目龐大的揮發性物理物質。許多情況下,受體-味道的結合是混雜的:也就是說,單一味道可能激活多個受體,而每位受體可能被多個味道激活。味覺神經元細胞各抒發一種味道受體。為此,每種味道都可以激活一組不同的(但有時是重疊的)神經元,繼而形成一個組合編碼,供神經系統破譯[3-5]。
果蠅和人類味道編碼的基本原則十分相像。諸如在這兩個物種中,抒發相同受體的味覺神經元的投射會聚在稱為嗅小球的樞紐狀結構處。但是,動物和腰部植物中的受體蛋白特別不同。肋骨植物味道受體是G蛋白偶聯受體家族的成員[6],而動物的受體是一些與味道分子結合時會打開的離子通道[7-10]。
幾六年來,這一現象人們早早已知曉,但尚不清楚單個味道受體怎樣對這么數目龐大結構不同的分子作出反應。1894年Emil首次提出受體-官能團結合的鎖鑰模型,該模型假設卟啉分子的形狀及其在受體中的結合位點完全互補[11]。但這不足以解釋受體的混雜性[12]。要闡明這一現象,須要了解與不同味道官能團結合的受體的結構。
為了實現這一目標,delMármol等人專注于動物味道受體。大多數這種受體由不同亞基(即異源多聚體)組裝而成,所有受體均包含一個特定亞基Orco和另一個賦于官能團特異性的可變亞基[13]。之前使用單粒子冷藏電子顯微鏡(cryo-EM)解析了包含四個Orco亞基的受體的結構[14],揭示了這些同源四聚體通道的基本結構。并且Orco不包含受體的官能團結合位點,因而結合位點仍未解析。為了解決動物味道受體的完整結構,同時規避解析異多聚體蛋白結構的困難,delMármol等人將眼神集中在一種不須要Orco即能組裝成功能性同源蛋白的動物味道受體上。
由相像亞基組成的異聚蛋白一般是在演變過程中基因復制時形成的。為此,delMármol等人推測在演進上更古老的生物體中會存在功能性同聚氣味受體,她們重點研究了蠹蟲的近親石蛃。在石蛃中,受體庫僅由五個亞基(MhOR1-5)組成,沒有一個與Orco直接相關[15]。每位MhOR5亞基包含九個α-螺旋,其中六個完全跨越細胞膜(S1-S6),另外兩個(S0和S7b)部份跨越細胞膜,最后一個(S7a)坐落細胞膜內。
作者測試了由四個MhOR5或四個MhOR1亞基組成的通道怎樣響應不同的官能團。她們發覺MhOR5通道被廣泛調諧細胞膜離子通道,對她們測試的60%以上的物理物質(包括丁香酚和避蚊胺)都有反應,而MhOR1通道更具選擇性。
接出來作者使用冷藏電鏡解析了處于非結合狀態(稱為apo狀態)、與避蚊胺、丁香酚結合時的MhOR5通道的結構(圖1)。總體而言,這種結構與Orco同聚體相像。
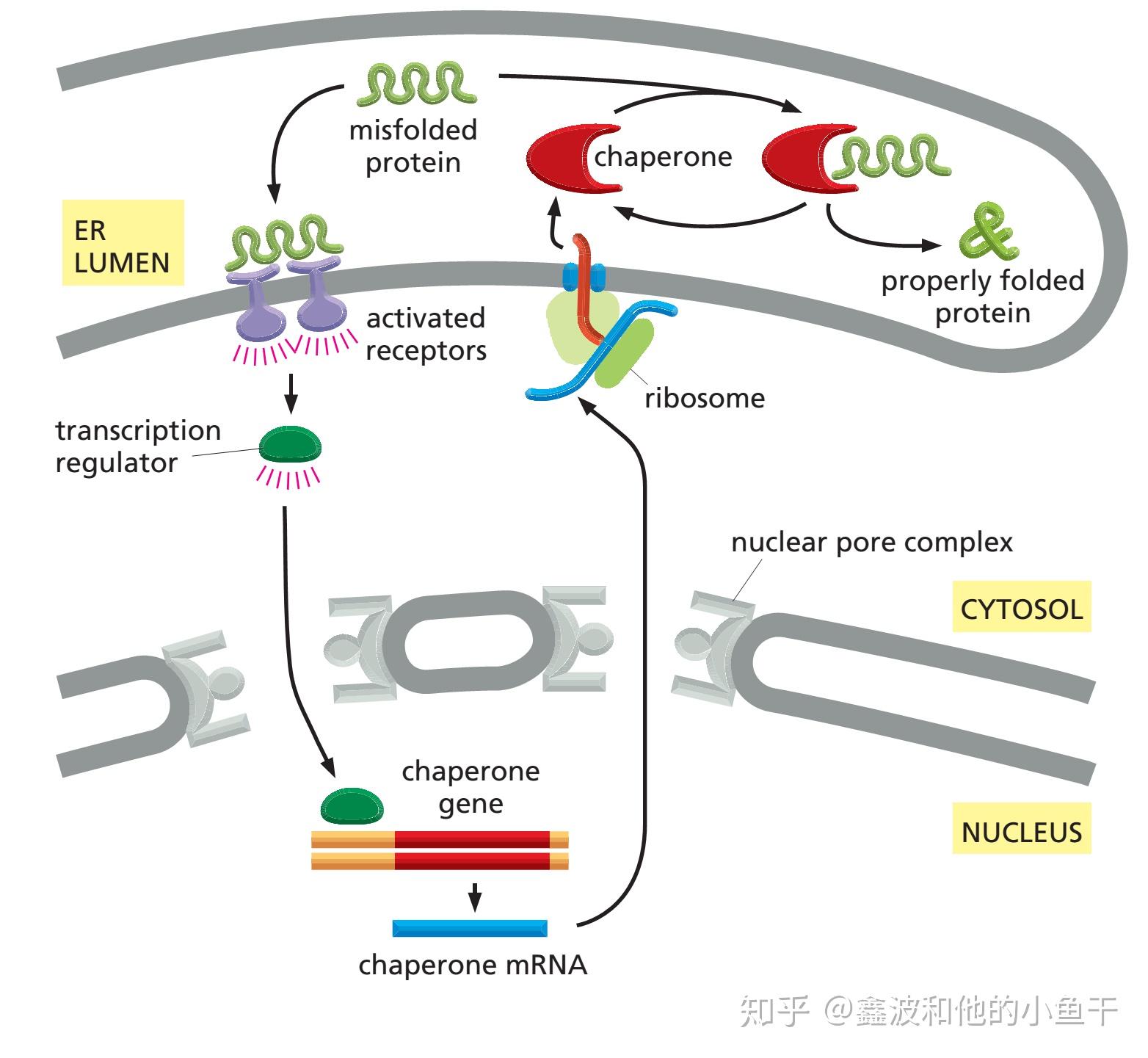
圖1|動物味道受體的閉合和打開狀態。
在動物中,味覺細胞上測量味道分子的受體是由四個蛋白質亞基組成的離子通道。delMármol等人[2]使用冷藏電子顯微鏡解析了來自石蛃()的一種味道受體處于非活性、未結合(apo)狀態及活性、開放狀態的結構。四個相同的亞基構成中心離子滲透通道;這兒分別展示了兩個亞基。當與丁香酚等味道劑結合時,每位亞基的S7b螺旋會輕微旋轉,進而擴大細胞膜外表面附近的通道孔。離子穿過一個大的前庭,之后通過四個縱向分支通道中的一個步入細胞。
這種結構表明,當通道被激活打開時,帶正電的離子順著一條通道從一個大的外前庭步入單個跨膜的通道,該通道可能是四個亞基中任意一個,均由S7bα-螺旋的多肽殘基排列而成。在受體的胞外側,四個通道從中心縱向分叉,構成離子步入細胞的通道的胞內部份。在未結合狀態下,該通道的最窄部份半徑為5.3埃,坐落S7b螺旋中的纈氨酸殘基塞入中央皺襞的位置細胞膜離子通道,產生疏水性“塞子”,限制離子通過。
未結合和官能團結合狀態的比較闡明了兩個關鍵的結構特點,這種特點其實能解釋味道受體通道一般怎樣才能對廣泛的官能團做出反應。首先,通道從關掉狀態到打開狀態的轉變僅涉及S7b螺旋的輕微旋轉,這會擴大孔徑并將極性殘基聯通到離子通道中。與在電流門控離子通道打開期間S4區域的運動相比,這些變化可能只須要極少的能量[16]。通道開啟所需能量這么之低,可能是味道受體官能團混雜的關鍵:雖然對受體具有低親和力的官能團的結合也會造成重排。
同樣重要的是卟啉結合位點的性質,它雖然異常靈活,才能在多個方面適應不同物理成份的絡合物。該絡合物結合口袋坐落由S2、S3、S4和S6螺旋的跨膜部份產生的松散束的深處,內襯主要是大的芳香族和疏水性殘基,這種殘基與官能團產生非極性互相作用。這與在許多其他類型受體的結合位點中觀察到的官能團(極性極強)嚴格的幾何限制產生鮮明對比。
據悉,MhOR5受體的結合位點恐怕會發生重排以適應不同大小和形狀的絡合物。口袋中的細微突變會改變官能團特異性,從本質上重新調整受體。這提供了一種機制,即進化可能會形成大量具有不同官能團特異性的受體,而這種受體在現存鳥類物種中早已被發覺。
總算,在發覺腰部植物味道受體的30年后[6],我們有了味道受體與其官能團復合物的結構。并且,雖然這一進展回答了一些古老的問題,它也提出了新的問題。諸如,不論是未結合的受體還是官能團結合受體的結構都沒有闡明味道劑步入官能團結合口袋的途徑。一種可能性是通道的“呼吸”,暫時打開一條通道,味道劑可以通過該通道到達該部位,進而將通道鎖定為開放構型。通過研究受體的結構和功能來定義味道通路可能會對受體特異性有的進一步了解。
這項研究和之前的研究也提出了一個問題,即激活狀態下的受體系統在沒有官能團的情況下(雖然處于低水平)怎樣分辨來自官能團結合受體的訊號和無官能團結合激活的干擾。一種可能性是味覺系統可以容忍一定量的干擾,由于它可以隨著時間的推移不斷搜集和整合新信息:比如,當一只動物緊靠味道源時,它可能會使用新的、更高質量的味覺信息來糾正它的路線。另一種可能性是觸覺系統組織成嗅小球,匯集來自許多神經元的信息,有效地過濾掉單個錯誤神經元的活動。可能須要剖析嗅小球和中樞神經系統對味道訊號的處理來回答那些問題。
最后,有人或許還想曉得這種發覺是否可以推廣到腰部植物G蛋白偶聯的味道受體。與動物中的味道受體一樣,腰部植物味道受體主要通過非極性和弱極性互相作用與味道結合[17]。它們是否也有靈活的官能團結合口袋和低能量屏障來參與下游訊號傳導,進而使它們還能對各類相對低親和力的官能團作出反應?毫無疑惑,delMármol等人的工作和未來的研究將指引為味道受體設計官能團的開發,其實可能有三天會有避蚊胺的取代品。