第二章細胞質膜
定義:圍繞在細胞最內層,由脂類、蛋白質和脂類組成的生物膜
意義:
1、在結構上作為細胞的界膜,使細胞具有一個相對穩定的內環境
2、在細胞與環境之間的物質運輸、能量轉換及信息傳遞的過程中起著重要的作用
生物膜:細胞內的膜系統與細胞質膜的總稱
第一節細胞質膜的結構模型與基本成份
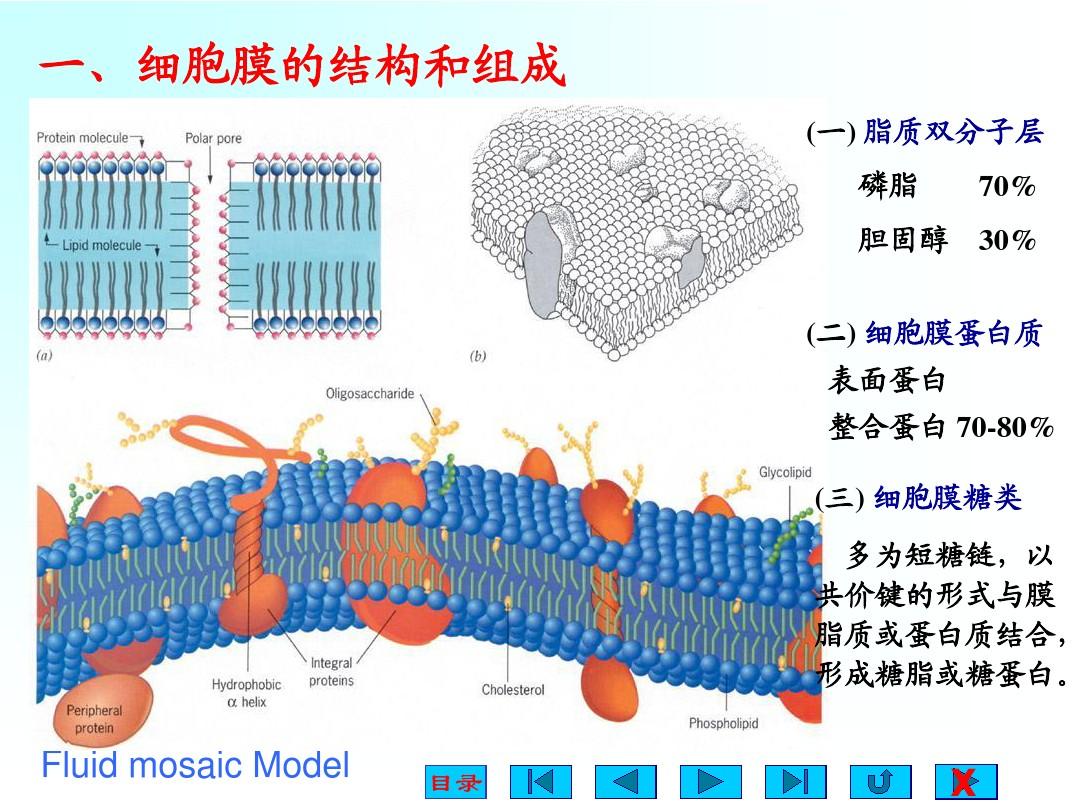
一、細胞質膜的結構模型
發展歷程:
1、人們利用光學顯微鏡觀察到了細胞,但始終沒有發覺細胞質膜
2、20世紀50年代初,電子顯微鏡發明,質膜的超微結構顯示下來(此時早已證明質膜的存在:滲透壓實驗)
3、1925年E.和F.用有機溶劑抽提了人的紅細胞質膜的膜脂成份,便于測定膜脂雙層分子在海面的描畫面積,發覺它是紅細胞表面積的二倍
4、和提出了“蛋白質-脂類-蛋白質”的披薩模型
5、1959年,J.D.依據電子顯微鏡觀察的結果提出了單位膜模型,發展了披薩模型,并大膽的推論所有的生物膜都由蛋白質-脂類-蛋白質的模型構成
6、細胞融合結合免疫螢光技術證明,質膜中的蛋白質是可流動的,電鏡冰凍刻蝕技術顯示了單層膜脂中富含蛋白質顆粒
7、1972年,S.J.和G.L.提出了流動鑲嵌模型,并得到各類實驗結果的支持,奠定了生物膜結構與特點的基礎
8、1975年,N.Unwin和R.首次報導了古核生物鹽病菌的質膜蛋白——菌紫色質的三維結構
9、脂筏模型提出:在甘油磷脂為主體的生物膜上,尿酸、鞘磷脂等富集區域產生相對有序的脂相,載著執行個別特定生物學功能的膜蛋白
流動鑲嵌模型的內容:
1、膜的流動性,即膜蛋白和膜質均可側向運動
2、膜蛋白分布的不對稱性,有的結合在膜表面,有的嵌入或橫越雙分子層
目前對生物膜結構的認知:
1、具有極性的腹部和非極性的尾部的磷脂分子在水相中具有自發產生封閉膜系統的性質,磷脂分子以疏水性尾部相對,極性的背部朝向水產生脂雙分子層,每層磷脂分子稱為一層小葉。脂分子是組成生物膜的基本結構成分,仍未發覺在生物膜結構中起組織作用的蛋白質。但在脂筏中存在某些有助于脂筏結構相對穩定的功能蛋白
2、蛋白質分子以不同的形式鑲嵌在脂雙分子層中或結合在其表面,蛋白質的類型,蛋白質分布的不對稱性及其與脂分子的協同作用賦于生物膜各自的特點和功能
3、生物膜可以看成是蛋白質在雙分子層的二維堿液,但是膜蛋白與膜脂之間、膜蛋白與膜蛋白之間及其與膜右側其他生物大分子的復雜的互相作用,在不同程度上限制了膜蛋白和膜脂的流動性。同時也產生了賴以完成多種膜功能的脂筏、纖毛和微絨毛等結構
4、在細胞生長和分裂等整個生命活動中,生物膜在三維空間上可出現彎曲、折疊、延伸等改變,處于不斷地動態變化中。因而保證了例如細胞運動、細胞增殖等代謝活動的進行
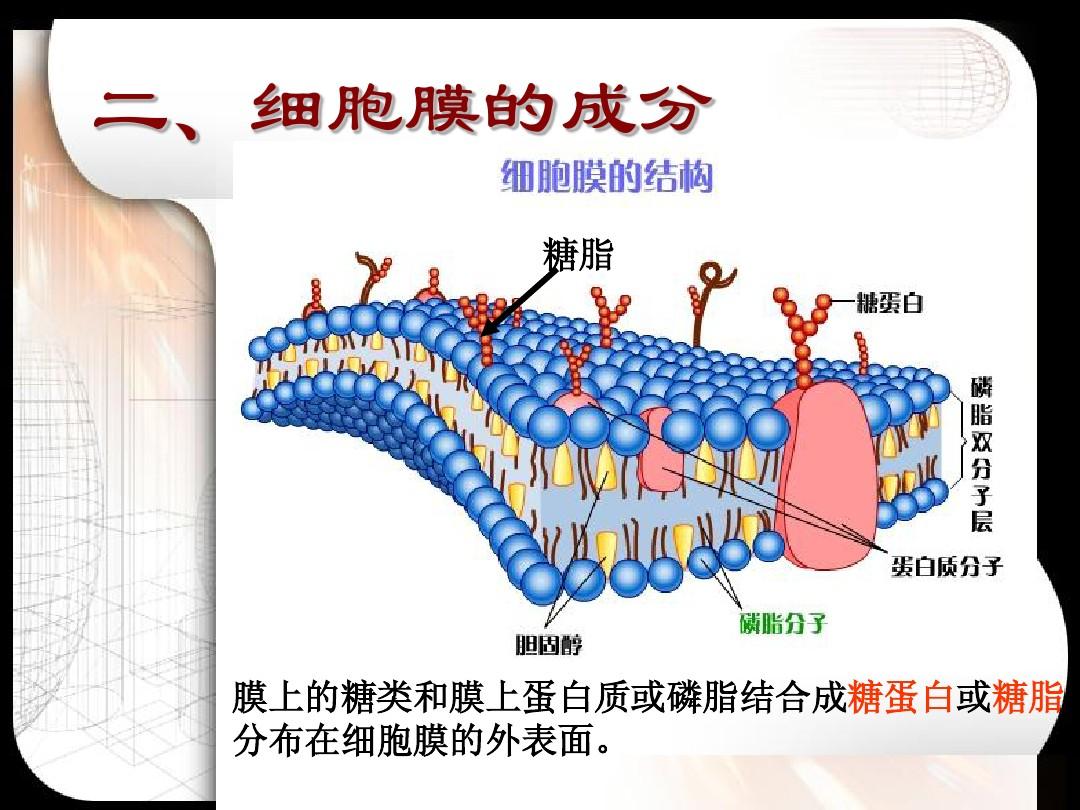
二、膜脂
生物膜的基本組成成份,每位植物細胞質膜約有109個脂分子,即每平方微米5*106個脂分子
(一)成份
主要包括甘油磷脂、鞘脂、固醇,以及少量的糖脂。鑒于大多數糖脂屬于鞘氨醇的衍生物,因而多將糖脂歸于鞘脂一類
1.甘油磷脂
構成膜脂的基本成份,占整個膜脂的50%以上。物理本質為3-乙酸甘油的衍生物,包括磷脂酰膽堿PC、磷脂酰丙氨酸PS、磷脂酰乙酸胺PE、磷脂酰肌醇PI。主要在葉綠體合成。
特點:
(1)具有一個與乙酸官能團相結合的極性頭和兩個非極性的尾(脂肪酸鏈),但存在于線粒體內膜和個別真菌質膜的心磷脂除外,它具有4個非極性尾部;極性頭的空間占位可影響脂單層的曲度,與PC相比,PE更容易產生曲鼻貼
(2)脂肪酸碳鏈為質數,多數碳鏈由16或18個碳原子組成
(3)除飽和脂肪酸(軟脂酸、硬脂酸)外,常富含1~2個苯環的不飽和脂肪酸(硬脂酸),不飽和脂肪酸多為順式,順式官能團在烴鏈中形成約30°角的彎曲
(4)甘油磷脂除了是生物膜的基本成份,但是其中的個別成份如PI等在細胞訊號轉導中起重要作用
2.鞘脂
均為鞘氨醇的衍生物,主要在高爾碳化物合成。具有一條烴鏈,另一條鏈是與鞘氨醇的羥基共價結合的長鏈脂肪酸。其腹部可能是一個類似于甘油磷脂的基于乙酸官能團的極性背部,稱為鞘磷脂SM,它的腹部是一個與鞘氨醇末端的甲基共價結合的乙酸膽堿。其分子結構與甘油磷脂十分相像,可以與甘油磷脂共同組成生物膜。鞘磷脂組成的脂雙分子層較甘油磷脂的寬度更大,如SM為4.6~5.6nm,而PC約3.5nm
糖脂:
兩性分子,它的極性背部直接共價結合到鞘氨醇上的一個分子或多糖鏈。糖脂普遍存在于原核和真核細胞的細胞質膜上,其濃度不足膜脂總數的5%,在神經細胞質膜上濃度較高,占5%~10%。
3.胰臟
固醇及其類似物也稱為嘌呤,它是一類富含四個閉環的碳溴化合物,其親水的一個背部為一個甲基,是一種分子剛性很強的兩性化合物
特征:
(1)與磷脂不同的是其分子的特殊結構和疏水性太強,自身不能產生脂雙分子層。只能插入磷脂分子之間,參與生物膜的產生
(2)固醇與甘油磷脂互相作用會降低磷脂分子的有序性及脂單層的長度,但對鞘磷脂沒有顯著的影響
(3)固醇存在于植物細胞和極少數的原核細胞中,在喂奶植物的細胞質膜中尤為豐富,其濃度通常不超過膜脂的1/3
(4)在多數細胞中,50%~90%的固醇存在于細胞質膜和相關的囊泡膜上
(5)固醇的合成是在植物細胞的胞質和葉綠體完成的,但動物體內的固醇多數來自于食物
(6)它在調節膜的流動性,降低膜的穩定性以及增加水溶性物質的私密性等起重要作用,是脂筏的基本結構
(7)缺少固醇可能造成細胞分裂的抑制
(8)動物細胞和細菌細胞都富含各自的嘌呤化合物,如動物中的豆甾醇和細菌中的麥角嘌呤。動物細胞質膜中的膽固醇濃度高達30%~50%,多數真菌質膜中不富含固醇成份,但個別真菌的膜脂中富含甘油酯等中性脂類
作用:
(1)重要的生物活性分子的前體化合物,如固丙酮激素、維生素D和膽酸等
(2)與發育調控的重要訊號分子共價結合
(3)通過濃度的調控控制生物膜的結構和功能
(二)膜質的運動形式
1.沿膜平面的側向運動氣溫為37℃時擴散系數為10-8cm2/s,相當于每秒聯通2μm的距離。因為側向運動形成分子間的換位,其交換頻度約為107次/s。側向運動是膜脂分子的基本運動形式,具有重要的生物學意義
2.脂分子圍繞軸心的載流子運動
3.脂分子尾部的擺動脂肪酸鏈緊靠腹部的擺動較小,其尾部的擺動較大。X射線衍射剖析顯示,在距腰部的第9個的碳原子之后的脂肪酸鏈已弄成無序狀態
4.單層脂分子之間的翻轉運動通常情況下翻轉運動很少發生,其發生的頻度還不到脂分子側向交換頻度的10-10。研究人工膜上脂分子運動所得到的結果與用原體、細菌和紅細胞膜為材料所得到的結果類似。但脂分子的翻轉運動在細胞個別膜系統中發生的頻度很高,非常是在核糖膜上,新合成的脂分子經幾分鐘后,將有半數從脂單層的右側通過翻轉運動轉位到另左側,這一過程須要特殊的膜蛋白完成
(三)脂類體
1.本質按照磷脂分子可在水相中產生穩定的脂單層膜的現象而制備的人工膜
2.產生雙層分子描畫在海面上時,其極性端插入水相而非極性端尾部面向空氣界面,攪亂后產生乳濁液,即產生極性端向外而非極性端向內的脂分子團或產生單層脂分子的球狀脂類體
3.特點球狀脂類體半徑為25~不等,控制產生條件可獲得大小均一的脂類體,同樣的原理還可以制備平面的脂類體膜
4.制備可用單一或混和的磷脂來制備,同時還可以嵌入不同的膜蛋白,因而脂類體是研究膜脂與膜蛋白及其生物學性質的極好的實驗材料
5.臨床診治脂類體與單克隆抗原等技術結合
三、膜蛋白
特征:
1.膜蛋白種類繁雜,酵母基因組中約1/3的基因編碼膜蛋白,多細胞有機體更多
2.多數膜蛋白的分子數目較少,但卻賦于生物膜十分重要的生物學作用
3.不同類型的細胞以及細胞不同部位的生物膜,其膜蛋白的濃度與種類都有很大區別
(一)膜蛋白的類型
基本分類:外在膜蛋白(外周膜蛋白)、內在膜蛋白(整合膜蛋白)、脂錨定膜蛋白
1.外在膜蛋白:
性質:水溶性蛋白質,靠離子鍵或其他較弱的鍵與膜表面的膜蛋白分子或膜脂分子結合
2.脂錨定蛋白
性質:通過與之共價相連的脂分子(脂肪酸或糖脂)插入膜的雙分子中,而錨定在細胞質膜上,其水溶性的蛋白質部份坐落脂單層外邊
分類:
(1)脂肪酸(軟脂酸)結合到膜蛋白N端的胺基殘基上
(2)由15到20個碳鏈長的內在膜蛋白烴鏈結合到膜蛋白C端的半胱谷氨酸殘基上,有時還有另一條烴鏈或脂肪酸鏈結合到近C端的其他半胱谷氨酸殘基上,這些雙重錨定有助于蛋白質更牢靠地與膜脂結合
(3)通過糖脂錨定在細胞質膜上
(二)內在膜蛋白與膜脂的結合形式
內在膜蛋白均為跨膜蛋白,跨膜蛋白在結構上可分為:胞質外結構域、跨膜結構域、胞質內結構域
與膜結合的主要方法:
1.膜蛋白的跨膜結構域與脂雙分子層中的疏水核心的互相作用,這是內在膜蛋白與膜脂結合的最主要和最基本的結合形式
2.跨膜結構域兩端攜帶正電荷的多肽殘基,如精谷氨酸、賴谷氨酸等與磷脂分子帶負電荷的極性臉部產生離子鍵,或帶負電荷的多肽殘基通過Ca2+、Mg2+等陽離子與帶負電的磷脂極性背部互相作用
3.個別膜蛋白通過自身在胞質兩側的半胱谷氨酸殘基共價結合到脂肪酸分子上,前者插入脂單層中進一步強化膜蛋白與脂單層的結合力
內在蛋白的跨膜結構域是與膜脂結合的主要部位,具體結構特征如下:
1.跨膜結構域富含20個左右的疏水多肽殘基,產生α螺旋(厚度約3nm),其外部疏水基團通過范德華力與脂雙分子層脂肪酸鏈(厚度約3.2nm)互相作用。這類蛋白也稱為單次跨膜蛋白(如血型糖蛋白A:跨膜α螺旋由23個多肽組成)。好多膜蛋白具有幾個跨膜的α螺旋區細胞膜選擇性通透,稱多次跨膜蛋白(G蛋白偶聯訊號通路的細胞表面受體:跨膜7次的膜蛋白)。跨膜結構域的α螺旋的方向,有的與膜面垂直細胞膜選擇性通透,有的則與膜面成一定角度。因而跨膜結構域的α螺旋寬度也各有差別
2.跨膜結構域主要由β折疊片組成(如大腸球菌外膜的孔蛋白,線粒體、葉綠體外膜上的孔蛋白)。因為α螺旋相鄰兩個多肽殘基的軸向距離為0.15nm,而在β折疊片中為0.35nm,所以跨膜結構域的β折疊片通常由10~12個多肽殘基組成。16個反向平行的β折疊片互相作用產生跨膜通道,通道具有疏水性的兩側和親水性的外側,可容許相對分子量大于104的小分子自由通過。孔蛋白跨膜結構域疏水性的兩側以及個別疏水性的基團使其穩定地結合在脂膜上。所有孔蛋白均為三聚體的跨膜蛋白
3.個別α羧基既具有極性基團又具有非極性基團。多個α螺旋產生特異極性分子的跨膜通道,其兩側為非極性鏈,與膜脂互相作用;外側是極性鏈,產生通道(如人紅細胞的帶3蛋白,介導Cl-和HCO3-的跨膜運輸)
*多數跨膜α螺旋的方向與脂膜平面成一定角度,一條最長的α螺旋其N端相對,各插進脂膜的一半,膜脂的種類和與膜蛋白的作用形式也直接影響到膜蛋白的空間構型及其功能
(三)去垢劑
本質:
是一端親水、一端疏水的兩性小分子,是分離與研究膜蛋白的常用試劑。去垢劑可以插入膜脂,與膜脂或膜蛋白的跨膜結構域等疏水部位結合,產生可溶的微粒
特征:
少量去垢劑能以單分子狀態溶化于水底,當達到一定含量時,去垢劑分子可在水底產生微團,此時去垢劑的含量稱為微團的臨界含量(CMC)。當使用的去垢劑含量低于或高于其CMC時,去垢劑的作用形式與膜蛋白的分離療效均有所不同。
分類:離子型去垢劑非離子型去垢劑
1.離子型去垢劑十二羥基氯化鈉SDS
具有帶電荷官能團
特征:
SDS可使細胞膜崩解,與膜蛋白疏水部份結合并使其與膜分離,高含量的SDS還可以破壞蛋白質中的離子鍵和官能團等非共價鍵,甚至改變蛋白質親水部份的構型
SDS對蛋白質的作用較為劇烈,可導致蛋白質變性,因而在純化膜蛋白時,非常是為獲得有生物活性的膜蛋白時,常采用不帶電荷的非離子型去垢劑
2.非離子型去垢劑X-100
非離子型去垢劑可使細胞膜崩解,但對蛋白質的作用比較溫和,它除了用于膜蛋白的分離與純化,也用于去除細胞的膜系統,便于對細胞骨架蛋白和其他蛋白質進行研究。
第二節細胞質膜的基本特點與功能
一、膜的流動性
意義:
1.膜的流動性是細胞質膜也是所有的生物膜的基本特點之一,也是細胞生長增殖等生命活動的必要條件
2.在脂膜二維空間上的熱運動是指膜脂和膜蛋白流動性的動力學基礎,膜脂與膜蛋白的互相作用以及與膜右側的生物大分子互相作用使膜的流動狀態更為復雜
3.除了保證了細胞正常的代謝活動并且受控于細胞代謝過程的調節
(一)膜脂的流動性
1.主要指脂分子的側向運動(由脂分子本身的性質決定)
影響誘因:
(1)通常來說,脂肪連越短,不飽和程度越高,膜脂的流動性越大
(2)氣溫對膜脂的運動有顯著的影響,各類膜脂都具有不同的相變體溫,鞘脂的相變氣溫通常低于磷脂
*在生物膜中膜脂的卜卦氣溫是由組成生物膜的各類脂分子的相變氣溫決定的(鞘脂或卵磷脂產生的脂單層膜流動性小一些,磷脂酰乙酸胺、磷脂酰肌醇和磷脂酰谷氨酸等產生的膜脂流動性大)
2.膜脂的流動性是生長細胞完成包括生長、增殖在內的多種生理功能所必需的,在真菌和植物細胞中經常通過降低不飽和脂肪酸的濃度來調節膜脂的相變氣溫以維持膜的流動
3.在植物細胞中,尿酸對莫得流動性也起著重要的雙重調節作用。固醇分子既有與磷脂疏水的尾部相結合使其更為有序、相互作用提高及限制其運動的作用,也有將磷脂分子隔開使其更易流動的功能。最終效應取決于尿酸在脂膜中的相對濃度以及上述兩種作用的綜合療效
4.因為膜脂與膜脂以及膜脂與膜蛋白之間的復雜的互相作用,膜質分子的運動狀態各不相同,其運動的區域也遭到一定的限制
(二)膜蛋白的流動性螢光抗原免疫標記實驗證明膜蛋白的流動性(人、鼠細胞雜交實驗)
現象:
1.膜蛋白在脂單層二維氨水中的運動是自發的熱運動,不須要細胞代謝產物的參與,也不須要能量輸出
2.在個別細胞中,當螢光抗原標記的時間繼續延長,已均勻分布在細胞表面的標記螢光會重新排布,集聚在細胞的個別部位(成斑現象),或集聚在細胞的一端(成帽現象)。解釋:二價的抗原分子交聯相鄰的膜蛋白分子,同時也與膜蛋白和膜下骨架系統的互相作用以及脂膜與細胞內膜系統之間的膜泡運輸有關
3.在極性細胞中,質膜蛋白被個別特殊的結構緊密聯接限定在細胞表面的某個區域。雖然在單細胞生物草履蟲的細胞質膜上,膜蛋白的分布也具有特定的區域性
4.用非離子去垢劑處理細胞使細胞膜系統崩解,多數膜蛋白流失,但仍有部份膜蛋白結合在細胞骨架上。細胞骨架不但影響膜蛋白的運動,也影響其周圍的膜脂的流動
5.膜蛋白與膜脂分子的互相作用也是影響膜流動性的重要誘因
(三)膜質和膜蛋白運動速度的測量
螢光漂白恢復(FPR)技術研究膜蛋白或膜脂流動性的基本實驗技術之一
原理:
用螢光素標記膜蛋白或膜脂,之后用激光束照射細胞表面某一區域,使被照射區的螢光淬滅變暗。因為膜的流動性,淬滅區域的色溫會漸漸降低,最后恢復到與周圍的螢光硬度相等。按照螢光恢復的速率可推斷出膜蛋白或膜脂的擴散速率
二、膜的不對稱性
膜脂和膜蛋白在生物膜上呈不對稱分布:同一種膜脂在脂雙分子層中的分布不同;同一種膜蛋白在脂單層中的分布都有特定的方向或拓撲學特點;糖蛋白和糖脂的糖基部份均坐落細胞質膜的內側
(一)細胞質膜各膜面的名稱
1.細胞外表面ES:與細胞外環境接觸的膜面,這一層脂分子和膜蛋白稱質膜的外小葉
2.原生質表面PS:與細胞質基質接觸的膜面,這一層脂分子和膜蛋白稱質膜的內小葉
3.細胞外小葉破裂面EF:電鏡冷藏刻蝕技術制樣過程中,膜結構往往從單層脂分子疏水端破裂,形成質膜的細胞外小葉破裂面
4.原生質小葉破裂面PF:電鏡冷藏刻蝕技術制樣過程中,膜結構往往從單層脂分子疏水端破裂,形成質膜的細胞內小葉破裂面
同理于細胞內的膜系統命名方法
*在脂肪細胞中和好多細胞中,都富含存儲脂肪的一種細胞器稱為脂滴,脂滴外圍僅有一層磷脂分子包被,相當于膜的內小葉。存儲脂肪在葉綠體膜的內外小葉之間合成,之后以發芽的方式披上葉綠體膜的內小葉,產生游離的脂滴。
(二)膜脂的不對稱性
指同一種膜脂分子在膜的脂單層中成不對稱分布。
1.多數磷脂存在于脂單層的內外兩邊,但某兩側常常濃度高一些,并非均勻分布
2.尿酸在生物膜內外小葉的分布通常比較均勻
3.糖脂的分布表現出完全的不對稱性,其糖基團都在質膜或在內膜的ES面上,因而糖脂僅存在于質膜的細胞外小葉中以的ES面。糖脂的不對稱分布是完成其生理功能的結構基礎
(三)膜蛋白的不對稱性
膜蛋白無論是在外在膜蛋白還是內在膜蛋白在質膜上都呈不對稱分布。與膜脂不同,膜蛋白的不對稱性是指每種膜蛋白分子在質膜上都有明晰的方向性
各類生物膜的特點及其生物學功能主要是由膜蛋白來決定。
膜蛋白的不對稱性是她們在合成時就早已決定,在此后的一系列轉運過程中其拓撲學結構一直保持不變直到蛋白質降解,而不會像膜脂那樣發生翻轉運動。
膜蛋白的不對稱性是生物膜完成復雜的在時間與空間上有序的各類生理功能的保障
三、細胞質膜相關的膜骨架
細胞質膜非常是膜蛋白往往與膜下結構(主要是細胞骨架系統)相互聯系、協同作用,并產生細胞表面的個別特化結構以完成特定的功能。這種特化結構包括膜骨架、鞭毛和肺臟、微絨毛及細胞的變型足等,分別與細胞形態的維持、細胞運動、細胞的物質交換和信息傳遞等功能有關。
(一)膜骨架
指細胞質膜下與膜蛋白相連的由纖維蛋白組成的網架結構,他從熱學上參與維持細胞質膜的形狀并協助質膜完成多種生理功能
(二)紅細胞的生物學特點
結構:紅細胞呈雙凹型的橢球結構,半徑約7μm,但它可以通過半徑比自己更小的毛細血管,平均壽命120天。喂奶植物成熟的紅細胞沒有細胞核和內膜系統
功能:把O2從肺運送到體內各組織,同時把細胞代謝形成的CO2運回肺中
血影:當紅細胞經低滲處理后,質膜斷裂,同時釋放流血紅蛋白和胞內其他可溶蛋白,此時剩下的紅細胞結構仍保持原先的基本形狀和大小,該結構稱為血影
(三)紅細胞質膜蛋白及膜骨架
紅細胞膜蛋白:血影蛋白(紅膜肽)、錨蛋白、帶3蛋白、帶4.1蛋白、帶4.2蛋白、肌動蛋白、血型糖蛋白
改變處理血影的離子硬度后進行電泳剖析,則血影蛋白和肌動蛋白條帶消失,說明這兩種蛋白質不是內在膜蛋白,比較容易去除。此時血影的形狀顯得不規則,膜蛋白的流動性提高,說明這兩種蛋白質在維持膜的形狀及固定其他膜蛋白的位置方面起重要作用
若用非離子型去垢劑X-100處理血影,這時帶3蛋白及一些血型糖蛋白的電泳條帶消失,但血影仍能保持原先的形狀,說明帶3蛋白及血型糖蛋白是膜整合蛋白,在維持血影乃至細胞形態上并不起決定性作用
紅細胞膜骨架的構成:
血影蛋白有α鏈和β鏈組成一個二聚體,長約100nm,半徑約5nm,兩個二聚體與腦部相連成一個寬度為200nm的四聚體,血影蛋白在體外氨水中就可以組裝。每位紅細胞約富含10萬個血影蛋白四聚體。與血影蛋白四聚體游離端相連的肌動蛋白纖維鏈長約35nm,其中包含13個肌動蛋白單體和一個原肌球蛋白分子(由兩個氨基酸組成3.5*104)。純化的血影蛋白與肌動蛋白纖維結合力十分微弱,帶4.1蛋白和一種稱為內收蛋白的蛋白質與之互相作用大大加大了肌動蛋白與血影蛋白的結合力。因為肌動蛋白纖維上存在多個與血影蛋白(通常為5個)結合位點,所以可以產生一個網路狀的膜骨架結構
膜骨架網路與細胞質膜之間的聯接主要通過錨蛋白。每位紅細胞中約富含10萬個錨蛋白分子。每位血影蛋白四聚體上平均有一個錨蛋白分子。錨蛋白富含兩個功能性結構域,一個能緊密地并且特異地與血影蛋白β鏈上的一個位點相連;另一個結構域與帶3蛋白中伸向胞質面的一個位點緊密結合,進而使血影蛋白網架與細胞質膜聯接在一起。據悉,帶4.1蛋白還可以與血型糖蛋白的細胞質結構域(C端)或帶3蛋白結合,同樣也起到使膜骨架與質膜蛋白相連的作用
*除紅細胞外,已發覺在其他細胞中也存在與錨蛋白、血影蛋白及帶4.1蛋白類似的蛋白質,大多數細胞中也存在膜骨架系統。與紅細胞不同,這種細胞具有較發達的胞質骨架系統,非常是質膜下呈網狀分布的肌動蛋白纖維,但是細胞質膜功能更為復雜。呈動態變化的膜骨架除了在熱學結構上為細胞質膜行使其功能提供一個三維的空間,并且直接參與細胞質膜的多種代謝活動。
(四)細胞質膜的基本功能
主要功能:
1.為細胞的生命活動提供相對穩定的內環境
2.選擇性的物質運輸,包括代謝底物的輸入與代謝產物的排除,其中伴隨著能量物質的傳遞
3.提供細胞辨識位點,并完成細胞內外信息跨膜傳導;病毒等病原微生物辨識和侵染特異的寄主細胞的受體也存在于質膜上
4.為多種酶提供結合位點,使酶促反應高效而有序地進行
5.介導細胞與細胞、細胞與胞外基質之間的聯接
6.質膜參與產生具有不同功能的細胞表面特化結構
7.膜蛋白的異常與個別遺傳病、惡性腦瘤、自身免疫病甚至神經退行性疾患相關,好多膜蛋白可作為癌癥醫治的抗生素靶標