《細(xì)胞膜電位》ppt講義恩格斯在100多年前總結(jié)自然科學(xué)成就時(shí)強(qiáng)調(diào):“地球幾乎沒有一種變化發(fā)生而不同時(shí)顯示出電的現(xiàn)象”;生物體其實(shí)也不例外。事實(shí)上,在希臘僅存史前古文字中,已有電魚擊人的記載;但對(duì)于生物電現(xiàn)象的研究,只能是在人類對(duì)于電現(xiàn)象通常規(guī)律和本質(zhì)有所認(rèn)識(shí)之后,并隨著電檢測(cè)儀器的精密化而日趨深入目前,對(duì)健康人和病人進(jìn)行心電圖、腦電圖、肌電圖,甚至黃斑電圖、胃腸電圖的檢測(cè),早已成為發(fā)覺、診斷和估量疾患進(jìn)程的重要手段;人體和各臟器的電現(xiàn)象的形成,是以細(xì)胞水平的生物電現(xiàn)象為基礎(chǔ)的,心電圖機(jī)腦電圖機(jī)肌電圖機(jī)黃斑電圖儀腸道電圖儀第1節(jié)剌激與反應(yīng)十九世紀(jì)中后期的生理學(xué)家用兩棲類植物做實(shí)驗(yàn)時(shí),發(fā)覺烏龜或蟾蜍的個(gè)別組織在離體的情況下,也能在一定的時(shí)間內(nèi)維持和表現(xiàn)出個(gè)別生命現(xiàn)象。這種生命現(xiàn)象的表現(xiàn)之一是:當(dāng)這種組織遭到一些外加的剌激誘因(如機(jī)械的、化學(xué)的、溫?zé)岬幕蜻m當(dāng)?shù)碾娯菁ぃ┳饔脮r(shí),可以應(yīng)答性出現(xiàn)一些特定的反應(yīng)或暫時(shí)性的功能改變。那些活組織或細(xì)胞對(duì)外界刺迸發(fā)生反應(yīng)的能力,就是生理學(xué)最早對(duì)于激動(dòng)性()的定義諸如,把蟾蜍的腓腸肌和支配它的神經(jīng)由體內(nèi)剝離下來,制成神經(jīng)-胸肌標(biāo)本,這時(shí)假若在神經(jīng)游離端兩側(cè)輕輕地打動(dòng)神經(jīng),或通以適當(dāng)?shù)碾妷海@么在經(jīng)過一個(gè)極短的潛伏期后,可以看見胸肌出現(xiàn)一次快速的減短和舒張;如把剌激直接施加于胸肌,也會(huì)造成類似的收縮反應(yīng);并且只要剌激不引起組織的損傷,上述反應(yīng)可以重復(fù)出現(xiàn)。
這就是神經(jīng)和胸肌組織具有激動(dòng)性能證明。實(shí)際上,幾乎所有活組織或細(xì)胞都具有某種程度的對(duì)外界刺迸發(fā)生反應(yīng)的能力,只是反應(yīng)的靈敏度和反應(yīng)的表現(xiàn)方式有所不同。在各類植物組織中,通常以神經(jīng)和肌細(xì)胞,以及個(gè)別腺細(xì)胞表現(xiàn)出較高的激動(dòng)性;這就是說它們只需接受較小的程度的剌激,才能表現(xiàn)出某種方式的反應(yīng),因而稱為可激動(dòng)細(xì)胞或可激動(dòng)組織。不同組織或細(xì)胞受剌激而發(fā)生反應(yīng)時(shí),外部可見的反應(yīng)方式有可能不同,如各類肌細(xì)胞表現(xiàn)機(jī)械收縮,腺細(xì)胞表現(xiàn)分泌活動(dòng)等,但所有那些變化都是由剌激造成的,因而把這種反應(yīng)稱之為激動(dòng)()。隨著電生理技術(shù)的發(fā)展和資料的積累,激動(dòng)性和激動(dòng)的概念有了新的涵義。大量事實(shí)表明,各類可激動(dòng)細(xì)胞處于亢奮狀態(tài)時(shí),盡管可能有不同的外部表現(xiàn),但它們都有一個(gè)共同的、最先出現(xiàn)的反應(yīng),這就是受剌激處的細(xì)胞膜外側(cè)出現(xiàn)一個(gè)特殊方式的電變化(它由細(xì)胞本身所形成,不應(yīng)與作為剌激使用的外加電剌激相混淆),這就是動(dòng)作電位;而各類細(xì)胞所表現(xiàn)的其他外部反應(yīng),如機(jī)械收縮和分泌活動(dòng)等,實(shí)際上都是由細(xì)胞膜的動(dòng)作電位進(jìn)一步觸發(fā)和引發(fā)的。在神經(jīng)細(xì)胞,非常是它的延續(xù)很長(zhǎng)、起著信息傳送作用的軸突(神經(jīng)纖維),在受剌激而激動(dòng)時(shí)并無肉眼可見的外部反應(yīng),其反應(yīng)只能用靈敏的電檢測(cè)儀器能夠測(cè)出的動(dòng)作電位。
在多數(shù)可激動(dòng)細(xì)胞(以神經(jīng)和骨骼肌、心肌細(xì)胞為主),當(dāng)動(dòng)作電位在受剌激部位形成后細(xì)胞膜電位,還可以順著細(xì)胞膜向周圍擴(kuò)布,使整個(gè)細(xì)胞膜都形成一次類似的電變化。神經(jīng)細(xì)胞示意圖既然動(dòng)作電位是大多數(shù)可激動(dòng)細(xì)胞受剌激時(shí)共有的特點(diǎn)性表現(xiàn),它不是細(xì)胞其他功能變化的伴隨物,而是細(xì)胞表現(xiàn)其他功能的前提或觸發(fā)誘因,因而在近代生理學(xué)中,激動(dòng)性被理解為細(xì)胞在受剌激時(shí)形成動(dòng)作電位的能力,而激動(dòng)一詞就成為形成動(dòng)作電位的過程或動(dòng)作電位的同義語了。只有這些在受剌激時(shí)能出現(xiàn)動(dòng)作電位的組織,能夠稱為可激動(dòng)組織;只有組織形成了動(dòng)作電位時(shí),才會(huì)說組織形成了激動(dòng)。可以對(duì)上述神經(jīng)-胸肌標(biāo)本的現(xiàn)象描述如下:當(dāng)剌激作用于坐骨神經(jīng)某一點(diǎn)時(shí),因?yàn)樯窠?jīng)纖維具有激動(dòng)性而出現(xiàn)激動(dòng),即形成了動(dòng)作電位,此動(dòng)作電位(常稱為神經(jīng)沖動(dòng))順著神經(jīng)纖維傳向它們所支配的骨骼肌纖維,通過神經(jīng)-肌接頭處的激動(dòng)傳遞,再造成骨骼肌細(xì)胞激動(dòng)而形成動(dòng)作電位,之后是動(dòng)作電位沿整個(gè)肌細(xì)胞膜傳遍整個(gè)肌細(xì)胞,并觸發(fā)了細(xì)胞內(nèi)收縮蛋白質(zhì)的互相作用,表現(xiàn)出胸肌一次快速的收縮和舒張。剌激導(dǎo)致激動(dòng)的條件和閾剌激具有激動(dòng)性的組織和細(xì)胞,并不對(duì)任何程度的剌激都能表現(xiàn)激動(dòng)或出現(xiàn)動(dòng)作電位。剌激可以泛稱細(xì)胞所處環(huán)境誘因的任何改變;泛指各類能量方式的理化誘因的改變,都可能對(duì)細(xì)胞構(gòu)成剌激。
在實(shí)驗(yàn)室中,常用各類方式的電剌激作為人工剌激,拿來觀察和剖析神經(jīng)或各類胸肌組織的激動(dòng)性,測(cè)度激動(dòng)性在不憐憫況下的改變。這是由于電剌激可以便捷地由各類電儀器(如電脈沖和方波發(fā)生器等)獲得,它們的硬度、作用時(shí)間和硬度-時(shí)間變化率可以容易地控制和改變;而且在通常情況下,就能造成組織激動(dòng)的電剌激并不導(dǎo)致組織損傷,因此可以重復(fù)使用。實(shí)驗(yàn)表明,剌激要造成組織細(xì)胞發(fā)生激動(dòng),必須在以下三個(gè)參數(shù)達(dá)到某一臨界值:剌激的硬度剌激的持續(xù)時(shí)間剌激硬度對(duì)于時(shí)間的變化率(即硬度對(duì)時(shí)間的微分)不僅這般,這三個(gè)參數(shù)對(duì)于導(dǎo)致某一組織和細(xì)胞的激動(dòng)并不是一個(gè)固定值,它們存在著互相影響的關(guān)系。為了說明剌激的各參數(shù)之間的互相關(guān)系,可以先將其中一個(gè)參數(shù)固定于某一數(shù)值,之后觀察其余兩個(gè)的互相影響。比如,當(dāng)使用方波剌激時(shí),因?yàn)椴煌笮『统掷m(xù)時(shí)間的方波上升支都以同樣極快的降低速度達(dá)到某一預(yù)定的硬度值,因此可以覺得上述第三個(gè)參數(shù)是固定不變的,而每一方波電剌激能夠?qū)е录?dòng),就只決定于它所達(dá)到的硬度和持續(xù)的時(shí)間了信噪比:在一串理想的脈沖周期序列中(如方波),正脈沖的持續(xù)時(shí)間與脈沖總周期的比值在神經(jīng)和肌組織進(jìn)行的實(shí)驗(yàn)表明,在硬度-時(shí)間變化率保持不變的情況下,在一定的范圍內(nèi),造成組織激動(dòng)所需的最小剌激硬度,與這一剌激所持續(xù)的時(shí)間呈反變的關(guān)系當(dāng)剌激的硬度較大時(shí),它只需持續(xù)較短的時(shí)間就足以引進(jìn)組織的激動(dòng),而當(dāng)剌激的硬度較弱時(shí),這個(gè)剌激就必須持續(xù)較長(zhǎng)的時(shí)間才會(huì)導(dǎo)致組織的激動(dòng)。
但這個(gè)關(guān)系只是當(dāng)所用硬度或時(shí)間在一定限度內(nèi)改變時(shí)是這么。假如將所用的剌激硬度降低到某一數(shù)值時(shí),則這個(gè)剌激不論持續(xù)多么長(zhǎng)也不會(huì)導(dǎo)致組織激動(dòng);與此相對(duì)應(yīng),假如剌激持續(xù)時(shí)間逐漸減短時(shí),最后也會(huì)達(dá)到一個(gè)臨界值,即在剌激持續(xù)時(shí)間大于這個(gè)值的情況下,無論使用多么大的硬度,也不能導(dǎo)致組織的激動(dòng)。假如用較小的剌激硬度能夠激動(dòng)的組織具有較高的激動(dòng)性,這么,這個(gè)硬度小的程度,還要決定于這個(gè)剌激的持續(xù)時(shí)間和它的硬度-時(shí)間變化率。為此,假如要簡(jiǎn)單地用剌激硬度這一個(gè)參數(shù)來表示不同組織激動(dòng)性的高低或同一組織激動(dòng)性的波動(dòng),就必須使所用剌激的持續(xù)時(shí)間和硬度-時(shí)間變化率固定為某一數(shù)值(應(yīng)是中等程度的);這樣,就能把造成組織激動(dòng)、即形成動(dòng)作電位所需的最小剌激硬度,作為評(píng)判組織激動(dòng)性高低的指標(biāo);這個(gè)剌激硬度稱為閾硬度或閾剌激,簡(jiǎn)稱閥值()。硬度大于閥值的剌激,稱為閾下剌激;閾下剌激不能導(dǎo)致激動(dòng)或動(dòng)作電位,但并非對(duì)組織細(xì)胞不形成任何影響。閥值越小,說明組織的激動(dòng)性越高。第2節(jié)細(xì)胞的靜息電位細(xì)胞水平的生物電現(xiàn)象主要有兩種表現(xiàn)方式,這就是它們?cè)诎察o時(shí)具有的靜息電位和它們?cè)獾截菁r(shí)形成的動(dòng)作電位。體內(nèi)各類臟器或多細(xì)胞結(jié)構(gòu)所表現(xiàn)的多種方式的生物電現(xiàn)象,大都可以按照細(xì)胞水平的那些基本電現(xiàn)象來解釋。
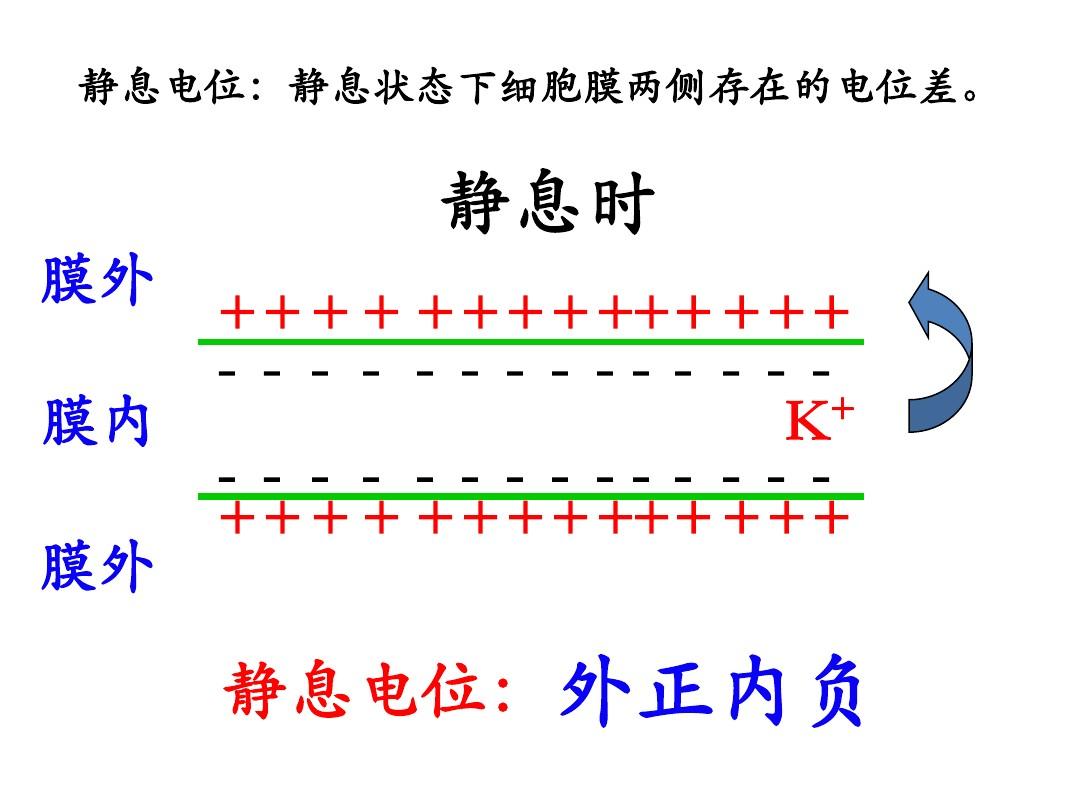
靜息電位指細(xì)胞未受剌激時(shí)存在于細(xì)胞內(nèi)外兩邊的電位差。當(dāng)兩個(gè)電極都處于膜外時(shí),只要細(xì)胞未遭到剌激或損傷,可發(fā)覺細(xì)胞外部表面各點(diǎn)都是等電位的;這就是說,在膜表面任意聯(lián)通兩個(gè)電極細(xì)胞膜電位,通常都不能測(cè)出它們之間有電位差存在。但若果讓微電極平緩地往前推動(dòng),讓它刺傷細(xì)胞膜步入膜內(nèi),這么在電極尖端剛才步入膜內(nèi)的頓時(shí),在記錄儀器中將顯示出一個(gè)忽然的電位躍變,這表明細(xì)胞膜內(nèi)外兩邊存在著電位差。K+Na+Cl-Na+Cl-K+膜內(nèi)膜外離子含量差電位差在靜息狀態(tài)下,細(xì)胞膜內(nèi)K+的高含量和安靜時(shí)膜主要對(duì)K+的私密性,是大多數(shù)細(xì)胞形成和維持靜息電位的主要誘因。(K+的平衡電位)膜外側(cè)的電-物理(含量)勢(shì)代數(shù)和為零時(shí),將不會(huì)再有K+的跨膜凈聯(lián)通K+多Cl-少Na+少內(nèi)負(fù)外正膜電位膜內(nèi)電位線性變化的一維膜模型dΦ(0)Φ(d)Intra--對(duì)于K+對(duì)上式從x=0到d積分Vm為跨膜電位,d為膜厚。設(shè)某種溶質(zhì)的脂水分配系數(shù)為k,其定義為:式中Cm及Cw分別代表溶質(zhì)在脂內(nèi)和在水底的溶化度。定義鉀的私密性Pk在細(xì)胞膜中,與鉀一起有著重要作用的還有鈉電壓JNa和氯電壓JCl。
總離子電壓為各組成部份之總和,即為:總的鉀離子通常而言,生物膜并不能讓所有的離子處于平衡中,假如對(duì)常見的組分估算K、Na、Cl的電位,其數(shù)值都不同。為此,靜息狀態(tài)僅僅能用穩(wěn)態(tài)來代表,即下述三種假設(shè)的條件下推論下來的:(1)如在堿液中一樣,膜內(nèi)離子也是在電場(chǎng)和含量梯度的影響下聯(lián)通的;(2)貼近膜的細(xì)胞內(nèi)離子含量和與其鄰接的氨水中的離子含量相等;(3)膜內(nèi)的電場(chǎng)梯度是均一的——方程/戈德曼多項(xiàng)式F是法拉第常數(shù),R是二氧化碳常數(shù),T是絕對(duì)濕度,P是通透常數(shù),e(extra)、i(intra)是細(xì)胞外,內(nèi)的離子含量。電位K+的電位烏賊神經(jīng)軸突細(xì)胞離子含量(mmol/L)細(xì)胞內(nèi)細(xì)胞外K++-+的電位為:檢測(cè)得到烏賊軸突細(xì)胞的靜息電位約為-70mV,因而在靜息時(shí)K+是接近(但不完全是)平衡的。1939年(霍奇金)等借助了槍烏賊的巨大神經(jīng)纖維和較精密的示波器等檢測(cè)儀器,第一次精確地測(cè)出此標(biāo)本的靜息電位值,結(jié)果發(fā)覺此值和估算所得的K+平衡電位值十分接近而略大于前者;如在一次實(shí)驗(yàn)中測(cè)得的靜息電位值為-77mV,而按當(dāng)時(shí)[K+]e和[K+]i值算出的Ek為-87mV。
大多數(shù)細(xì)胞的靜息電位的形成,是因?yàn)檎<?xì)胞的細(xì)胞內(nèi)液高K+而膜在安靜時(shí)又主要對(duì)K+有通透能力的結(jié)果;至于靜息電位的數(shù)值為什么略大于理論上的Ek值,通常覺得是因?yàn)槟ぴ陟o息時(shí)對(duì)Na+也有極小的私密性(大概只有K+私密性的1/50~1/100)的緣故;因?yàn)槟ね釴a+含量小于膜內(nèi),雖然小量的Na+逸入膜內(nèi)也會(huì)抵消一部份K+外移導(dǎo)致的膜內(nèi)負(fù)電位。第3節(jié)神經(jīng)細(xì)胞神經(jīng)元也叫神經(jīng)細(xì)胞,是構(gòu)成神經(jīng)系統(tǒng)結(jié)構(gòu)的基本單位。神經(jīng)元是具有長(zhǎng)凸出的細(xì)胞,它由細(xì)胞體和細(xì)胞凸出構(gòu)成。細(xì)胞體坐落腦、脊髓和神經(jīng)節(jié)中,細(xì)胞凸出可延展至腰部各臟器和組織中。細(xì)胞體是細(xì)胞含核的部份,其形狀大小有很大差距,半徑約4~120微米。核大而圓,坐落細(xì)胞中央,染色質(zhì)少,胚乳顯著。細(xì)胞質(zhì)內(nèi)有斑片狀的核外染色質(zhì)(舊稱尼爾小體),還有許多神經(jīng)元纖維。細(xì)胞凸起是由細(xì)胞體延展下來的狹長(zhǎng)部份,又可分為樹突和軸突。每位神經(jīng)元可以有一或多個(gè)樹突,可以接受剌激并將激動(dòng)傳入細(xì)胞體。每位神經(jīng)元只有一個(gè)軸突,可以把激動(dòng)從胞體傳送到另一個(gè)神經(jīng)元或其他組織,如胸肌或胃壁。在外周神經(jīng)系統(tǒng)中,我們所見到的神經(jīng)纖維都是軸突。各類神經(jīng)元軸突粗細(xì)長(zhǎng)短均不相同,通常較粗的軸突傳導(dǎo)速率較快,反之較慢絕大多數(shù)軸突半徑為30~50微米。
在一些大動(dòng)物體內(nèi)軸突可以歷時(shí)幾米,人體中最長(zhǎng)的軸突大概為一米。在電生理實(shí)驗(yàn)中,可以以一定硬度與頻度的電壓脈沖剌激細(xì)胞,就形成了動(dòng)作電位。通常的實(shí)驗(yàn)對(duì)象是馬蹄蟹的視神經(jīng)纖維或?yàn)踬\魚巨纖維做實(shí)驗(yàn)。借助紅蟲神經(jīng)索的巨纖維也可挺好地說明在單根纖維上傳導(dǎo)著的動(dòng)作電位的行為。第3節(jié)動(dòng)作電位指可激動(dòng)細(xì)胞遭到剌激而激動(dòng)時(shí),在靜息電位的基礎(chǔ)上膜外側(cè)的電位發(fā)生快速而可逆的倒轉(zhuǎn)和復(fù)原。這些電位變化叫做動(dòng)作電位極化()——膜外側(cè)存在的內(nèi)負(fù)外正的電位狀態(tài)。去極化()——膜電位絕對(duì)值漸漸減弱的過程。超極化(Over-)——膜電位絕對(duì)值低于靜息電位的狀態(tài)。復(fù)極化()——膜電位去極化后逐漸恢復(fù)極化狀態(tài)的過程。動(dòng)作電位,相對(duì)于靜息電位,是細(xì)胞和組織去極化的過程。它是具有一定頻度和振幅特點(diǎn)的生物電位,是活系統(tǒng)激動(dòng)的一種外在表現(xiàn)。動(dòng)作電位的出現(xiàn)是一切激動(dòng)細(xì)胞和組織,非常是比較高等、復(fù)雜結(jié)構(gòu)生物對(duì)象的本性。它可起因于組織本身的內(nèi)在過程,如腦電、心電等,也可來自外界的人為形成,如離體細(xì)胞組織在不同剌激誘因作用下形成的電位,或來自外部體會(huì)裝置的傳入脈沖。
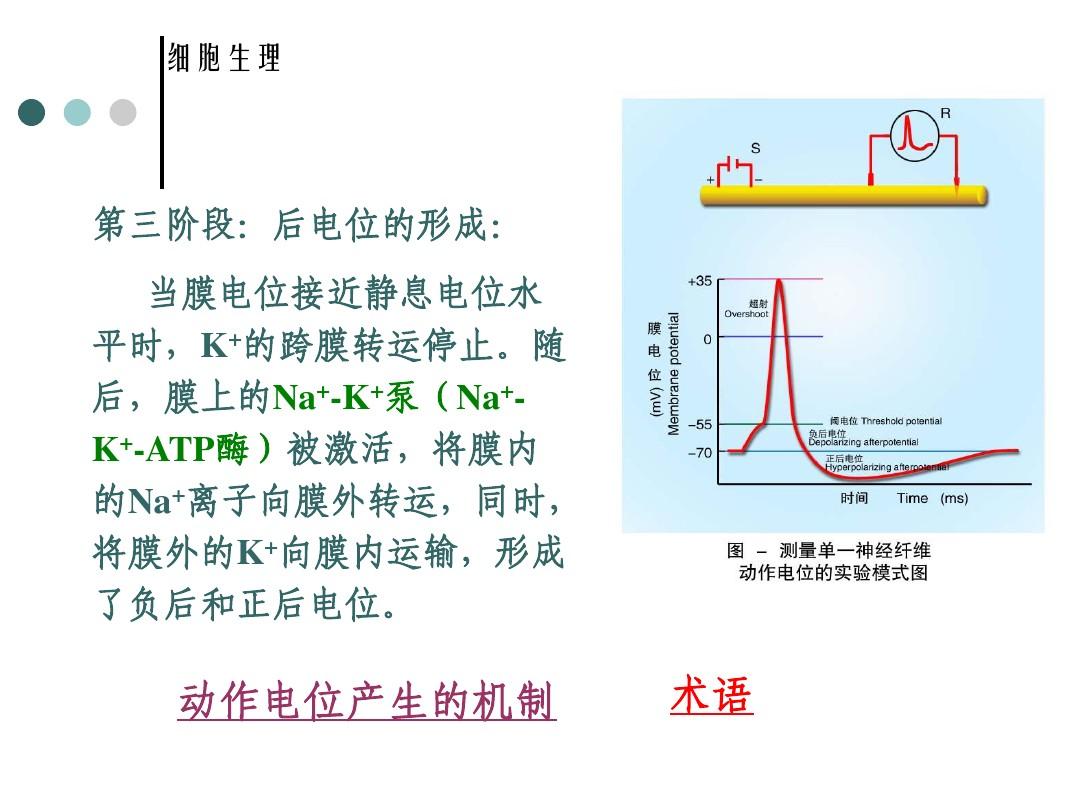
動(dòng)作電位包括三個(gè)基本過程:(1)去極化,膜內(nèi)原先存在的負(fù)電位迅速消失,即膜電位的極化狀態(tài)消失。(2)超極化,繼去極化以后,繼而發(fā)展為極化狀態(tài)倒轉(zhuǎn),即轉(zhuǎn)變?yōu)槟?nèi)為正,膜外為負(fù)。(3)復(fù)極化,膜內(nèi)電位達(dá)到頂峰后開始回升,恢復(fù)至原先靜息電位水平。第一階段:動(dòng)作電位上升支的產(chǎn)生(去極化相的產(chǎn)生)形成緣由:因?yàn)樨菁ぴ斐赡?duì)Na+的私密性頓時(shí)減小(Na離子通道被激活),膜外的Na+內(nèi)流,使膜電位由-70mV降低至0mV,因而上升為+30mV,Na+通道急劇失活。第二階段:動(dòng)作電位升高支產(chǎn)生:Na+通道失活后,膜恢復(fù)了對(duì)K+的私密性,大量的K+外流。使膜電位由正值向負(fù)值轉(zhuǎn)變,產(chǎn)生了動(dòng)作電位的增長(zhǎng)支。動(dòng)作電位是在極短的時(shí)間內(nèi)形成的,因而,在體外描記的圖形為一個(gè)短促而尖銳的脈沖圖形,似山峰般,稱為峰電位()。第三階段:后電位的產(chǎn)生:當(dāng)膜電位接近靜息電位水平時(shí),K+的跨膜轉(zhuǎn)運(yùn)停止。此后,膜上的Na+-K+泵(Na+-K+-ATP酶)被激活,將膜內(nèi)的Na+離子向膜外轉(zhuǎn)運(yùn),同時(shí),將膜外的K+向膜內(nèi)運(yùn)輸,產(chǎn)生了負(fù)后和正后電位。通過解釋靜息電位和峰值電位的來源早已初步剖析了動(dòng)作電位波形的主要方面,并且要想考慮動(dòng)作電位的整個(gè)時(shí)間過程,這種還是不夠的。
比如為何電位上升很快而回降較慢?當(dāng)考察同一種昆蟲的不同神經(jīng)和胸肌上的動(dòng)作電位時(shí),或是當(dāng)考察不同種鳥類的同一神經(jīng)或胸肌上的動(dòng)作電位時(shí),為什么觀察到在動(dòng)作電位的形狀和時(shí)間過程上有許多變化等等。要想回答有關(guān)時(shí)間過程的問題就須要有關(guān)膜行為的更適宜的模型,而這必須構(gòu)建在廣泛的實(shí)驗(yàn)結(jié)果的基礎(chǔ)上。1.膜的熱學(xué)模型和并聯(lián)濁度模型——這個(gè)模型給出了剖析各個(gè)組成離子電壓的框架2.電流嵌位實(shí)驗(yàn)的結(jié)果——電壓嵌位實(shí)驗(yàn)用以評(píng)價(jià)各個(gè)離子電壓的最重要的實(shí)驗(yàn)工具3.-等式。這種等式總結(jié)了由電流嵌位實(shí)驗(yàn)所獲得的實(shí)驗(yàn)數(shù)據(jù),成功地解釋了動(dòng)作電位的各個(gè)方面,描述了相應(yīng)的各類離子的運(yùn)動(dòng)。4.借助-多項(xiàng)式進(jìn)行動(dòng)作電位的數(shù)值仿真。第4節(jié)細(xì)胞膜的熱學(xué)模型第5節(jié)電流固定的膜電壓研究第6節(jié)-多項(xiàng)式第7節(jié)對(duì)膜動(dòng)作電位的仿真膜電位/靜息電位:細(xì)胞生命活動(dòng)過程中伴隨的電現(xiàn)象,存在于細(xì)胞膜外側(cè)的電位差稱膜電位。()一般是指以膜相隔的兩堿液之間形成的電位差。生物細(xì)胞被以半透性細(xì)胞膜,而膜兩側(cè)呈現(xiàn)的生物電位就是這些電位,平時(shí)把細(xì)胞內(nèi)外的電位差叫膜電位。
假如把兩種電解質(zhì)用膜隔開,使左側(cè)富含不能透過該膜的粒子,因?yàn)檫@些影響,左側(cè)電解質(zhì)的分布便發(fā)生了變化,一旦董南()膜平衡完善膜右側(cè)還會(huì)有董南膜電位。假如兩邊沒有這些不透性離子,但只要把含量不同的兩種電解質(zhì)以膜隔開,在陽離子和陰離子透過膜的速率不同時(shí),膜外側(cè)也會(huì)形成電位差。在膜右側(cè)放0.1和0.01N的KCl堿液時(shí)形成的膜電位,作為表現(xiàn)膜特點(diǎn)的電位,則稱為標(biāo)準(zhǔn)電位差,其值最大可達(dá)58mV。膜電位的存在和各類影響導(dǎo)致的這種變化是靜止電位和動(dòng)作電位的動(dòng)因。閾電位:當(dāng)細(xì)胞遭到一次閾剌激或閾上剌激時(shí),受激細(xì)胞膜上Na通道少量開放,出現(xiàn)Na少量?jī)?nèi)流,使膜的靜息電位值減少而發(fā)生去極化。當(dāng)去極化進(jìn)行到某一臨界值時(shí),因?yàn)镹a通道的電流依從性,導(dǎo)致Na通道大量激活、開放,致使Na迅速大量?jī)?nèi)流而爆發(fā)動(dòng)作電位。這個(gè)足以使膜上Na通道忽然大量開放的臨界膜電位值,稱為閾電位。閾電位比靜息電位約小10mV~20mV。如神經(jīng)纖維的靜息電位是-70mV,其閾電位約為-55mV。任何剌激只要能使膜從靜息電位去極化到閾電位,便能觸發(fā)動(dòng)作電位,導(dǎo)致激動(dòng)。有人將閾電位稱為燃點(diǎn),這是十分形象化的術(shù)語。從電生理的角度來看,激動(dòng)是指動(dòng)作電位的形成過程或動(dòng)作電位的同義語,而激動(dòng)性則是細(xì)胞受剌激時(shí)形成動(dòng)作電位的能力。
激動(dòng)性的基礎(chǔ)是靜息電位,所以靜息電位值或靜息電位與閾電位的距離大小,可影響細(xì)胞的激動(dòng)性。如二者距離減小,細(xì)胞的激動(dòng)性增長(zhǎng)。動(dòng)作電位(1)概念:可激動(dòng)組織或細(xì)胞遭到閾上剌激時(shí),在靜息電位基礎(chǔ)上發(fā)生的快速、可逆轉(zhuǎn)、可傳播的細(xì)胞膜外側(cè)的電變化。動(dòng)作電位的主要成份是峰電位。(2)產(chǎn)生條件:①細(xì)胞膜外側(cè)存在離子含量差,細(xì)胞膜內(nèi)K+含量低于細(xì)胞膜外,而細(xì)胞外Na+、Ca2+、Cl-低于細(xì)胞內(nèi),這些含量差的維持借助離子泵的主動(dòng)轉(zhuǎn)運(yùn)。(主要是Na+-K+泵的轉(zhuǎn)運(yùn))。②細(xì)胞膜在不同狀態(tài)下對(duì)不同離子的私密性不同,比如,安靜時(shí)主要容許K+通透,而去極化到閾電位水平時(shí)又主要容許Na+通透。③可激動(dòng)組織或細(xì)胞受閾上剌激。(3)產(chǎn)生過程:≥閾剌激→細(xì)胞部份去極化→Na+少量?jī)?nèi)流→去極化至閾電位水平→Na+內(nèi)流與去極化產(chǎn)生正反饋(Na+爆發(fā)性內(nèi)流)→達(dá)到Na+平衡電位(膜內(nèi)為正膜外為負(fù))→形成動(dòng)作電位上升支。膜去極化達(dá)一定電位水平→Na+內(nèi)流停止、K+迅速外流→形成動(dòng)作電位升高支。(4)產(chǎn)生機(jī)制:動(dòng)作電位上升支——Na+內(nèi)流所致。動(dòng)作電位的幅度決定于細(xì)胞內(nèi)外的Na+含量差,細(xì)胞外液Na+含量減少動(dòng)作電位幅度也相應(yīng)增加,而阻斷Na+通道(河豚毒)則能制約動(dòng)作電位的形成。
動(dòng)作電位升高支——K+外流所致。動(dòng)作電位時(shí)細(xì)胞遭到剌激時(shí)細(xì)胞膜形成的一次可逆的、可傳導(dǎo)的電位變化。形成的機(jī)制為①閾剌激或閾上剌激使膜對(duì)Na+的私密性降低,Na+順含量梯度及電位差內(nèi)流,使膜去極化,產(chǎn)生動(dòng)作電位的上升支。②Na+通道失活,而K+通道開放,K+外流,復(fù)極化產(chǎn)生動(dòng)作電位的增長(zhǎng)支。③鈉泵的作用,將步入膜內(nèi)的Na+泵出膜外,同時(shí)將膜外多余的K+泵入膜內(nèi),恢復(fù)激動(dòng)前是離子分布的含量。神經(jīng)膜的并聯(lián)濁度模型給出了一小塊神經(jīng)膜的一個(gè)簡(jiǎn)單的熱學(xué)表示,它被稱為并聯(lián)濁度模型。每一支路表示某一特定種類的離午時(shí)整個(gè)跨膜電壓的貢獻(xiàn)。假定對(duì)K,Na,Cl有獨(dú)立的濁度通道。細(xì)胞外細(xì)胞內(nèi)第4節(jié)細(xì)胞膜的熱學(xué)模型內(nèi)負(fù)外正膜電位K+多Cl-少Na+少InOut舉例說,假如膜電位是Vm,那末鉀的凈驅(qū)動(dòng)力是(Vm-EK),也就是對(duì)平衡條件的偏離。由于鉀電壓反比于電流(Vm-EK),因而比列系數(shù)就是濁度的單位。把鉀濁度記為gK(其值依賴于Vm和t),因而靜息時(shí),Ic=0要想列舉對(duì)跨膜電壓的所有貢獻(xiàn),我們還要加上電容性(或位移)電壓,它就是穩(wěn)態(tài)時(shí),這就是并聯(lián)濁度多項(xiàng)式。它描述了Vm如何作為與相對(duì)導(dǎo)電性有關(guān)的EkECl和ENa的某種加權(quán)平均。
這一表達(dá)式只有在假設(shè)的穩(wěn)態(tài)條件下才是正確的。由烏賊軸突的細(xì)胞內(nèi)外各類離子的含量值,我們可到其電位:Ek=-74.=-65.=54.2mV假設(shè)存在下述“典型”值:gk=0.367mS/=0.582mS/=0.010mS/cm2我們來看電位和導(dǎo)電性對(duì)穩(wěn)態(tài)跨膜電位有何影響。由以上數(shù)據(jù)代入式并聯(lián)濁度多項(xiàng)式,得到Vm=-68.0mV對(duì)于這樣的靜息電位才會(huì)有穩(wěn)定的鉀外流。這個(gè)外流為Vm和Ek差值6.7mV所驅(qū)動(dòng)。這時(shí)也會(huì)有鈉的內(nèi)流,它為Vm偏離開它的平衡電位的差值122.2mV所驅(qū)動(dòng)。這個(gè)大的驅(qū)動(dòng)力作用在比較低的導(dǎo)電性上,因而鉀的外流和鈉的內(nèi)流大致平衡而維持穩(wěn)態(tài)(我們?nèi)匀患僭O(shè)氯實(shí)際上處于平衡)。Ek=-74.=-65.=54.2mV第5節(jié)電流固定的膜電壓研究“電壓嵌位”一直是用以評(píng)估動(dòng)作電位期間離子電壓時(shí)間過程的最重要的工具。電流嵌位追溯在動(dòng)作電位期間,跨膜電壓的成份包括離子流和電容性(充電)電壓。由于膜電容是不變的,則前者等于CmdVm/dt。若果在膜兩側(cè)加上前饋電流(即加上恒定的跨膜電位),電路中就不再有電容性成份,因此也就簡(jiǎn)化了對(duì)有關(guān)電壓的剖析,這時(shí)電壓必將完全由離子成份構(gòu)成。
由歐姆定理可知,內(nèi)阻一定時(shí),電壓發(fā)生改變,必然導(dǎo)致膜電位急劇變化,這樣就難以觀察膜電位對(duì)離子流的影響。通過將膜電位鉗制在不同水平,以防止離子流反過來影響電流值。通常而言,膜對(duì)某種離子私密性的變化是膜電位和時(shí)間的函數(shù)。用玻璃微電極插入細(xì)胞內(nèi),借助電子學(xué)技術(shù)施加一跨膜電流并把膜電位固定于某一數(shù)值,可以測(cè)定該膜電位條件下離子電壓隨時(shí)間變化的動(dòng)態(tài)過程。借助抗生素或改變細(xì)胞內(nèi)外的氨水成份,使其他離子通道失效,即可測(cè)定被研究的某種離子通道的功能性熱阻,剖析離子電壓的穩(wěn)態(tài)和動(dòng)力學(xué)與膜電位、離子含量等之間的關(guān)系,可推測(cè)該種通道的濁度、活化和失活速度、離子選擇性等,并能檢測(cè)和剖析通道的門控電壓的特點(diǎn)。圖中電極1插入巨大神經(jīng)軸突內(nèi)一定距離,拿來檢測(cè)和監(jiān)察這一段軸突膜內(nèi)的電位,此電極先連到一個(gè)電流放大器,再在一個(gè)示波器上顯示;電極1測(cè)得電位值經(jīng)放大后同時(shí)輸給一個(gè)負(fù)反饋放大器(FBA),這是整個(gè)儀器設(shè)計(jì)的關(guān)鍵部份,它可把測(cè)得的膜內(nèi)電位同來自一個(gè)電流源的、由實(shí)驗(yàn)者預(yù)先設(shè)定的要求保持恒定的電位值進(jìn)行比較,假如兩者有差值,F(xiàn)BA都會(huì)通過電極2向軸突膜內(nèi)輸出相應(yīng)硬度和方向的電壓,因?yàn)閮x器線路的精密設(shè)計(jì)和快速反應(yīng),電極2輸出電壓的改變正足以補(bǔ)償標(biāo)本因?yàn)榭缒るx子電壓使膜充放電而導(dǎo)致的跨膜電位的變動(dòng),于是與電極1相邊的示波器上顯示出膜內(nèi)電位固定在設(shè)定的數(shù)值,而在電流放大器IA上測(cè)得的跨膜離子電壓的變化,就反映了膜濁度的變化。
020-(ms)(mA/cm2)al(mV)在t=0時(shí)應(yīng)用電流嵌位條件下烏賊軸突的離子電壓Vm=20mV,靜息電位-(mA/cm2)在t=0時(shí)應(yīng)用電流嵌位條件下烏賊軸突的離子電壓,靜息電位-60mV,ENa=57mV能斯特電位電流鉗實(shí)驗(yàn)結(jié)果示意圖單獨(dú)觀察Na+電壓,可用TEA(,四甲基胺)阻斷K+外流后得到;單獨(dú)觀察K+外流,則用TTX(,河豚毒)阻斷Na+內(nèi)流后得到第6節(jié)-多項(xiàng)式(霍奇金)和(赫胥黎)從她們?cè)跒踬\軸突的電流嵌位實(shí)驗(yàn)中所收集得到的數(shù)據(jù)構(gòu)建了知名的定量模型。模擬電流嵌位實(shí)驗(yàn)數(shù)據(jù)模擬動(dòng)作電位傳播將離子電壓和其驅(qū)動(dòng)力聯(lián)系上去濁度的計(jì)算方式:按照相應(yīng)的電流嵌實(shí)驗(yàn)數(shù)據(jù),換算出相應(yīng)的濁度數(shù)據(jù)關(guān)于鉀濁度的假設(shè):關(guān)于鈉濁度:-方程式中的n、m和h都是機(jī)率;第7節(jié)對(duì)膜動(dòng)作電位的仿真采用-多項(xiàng)式對(duì)膜動(dòng)作電位進(jìn)行仿真假設(shè)開始時(shí)在一段時(shí)間T內(nèi)的去極化電壓為Id把時(shí)間t離散化-~dst/HHsim/