核被膜
中文名稱:
真核生物的細胞核的最內層結構,由兩層單位膜所組成。它將DNA與細胞質隔開,產生了核內特殊的微環境,保護DNA分子免受損傷;使DNA的復制和RNA的翻譯抒發在時空上分隔開來;據悉染色體定位于核膜上,有利于解旋、復制、凝縮、平均分配到子核,核被膜還是核質物質交換的通道。
目錄
核被膜的結構
核被膜由內核膜(inner)、外核膜(outer)和核周隙(space)三部份構成。核被膜上有核孔與細胞質相通。
核被膜()包裹在核表面,由基本平行的外層膜、外層膜兩構成。兩層膜的間隙寬10~15nm,稱為核周隙()。核被膜上有核孔(pore)穿通。外核膜表面有內質網體附著,并與粗面核糖相續;核周隙亦與核糖腔相通,為此,核被膜也參與蛋白質合成。內核膜也參與蛋白質合成。內核膜的核質面有厚20~80nm的核纖層(),是一層由細絲交織產生的致密網狀結構。核纖層除了對核膜有支持、穩定作用,也是染色質纖維西端的附著部位。
一、核纖層定義
外核膜胞質面附有內質網體,并與核糖相連,核周隙與核糖腔相通,可以說是葉綠體的一部份。外核膜上附著10nm的中間纖維(),可見核是被核糖和中間纖維相對固定的。
核周隙寬20~40nm,腔內電子密度低,通常不含固定的結構。
內核膜的內表面有一層網路狀纖維蛋白質,叫核纖層(),可支持核膜。
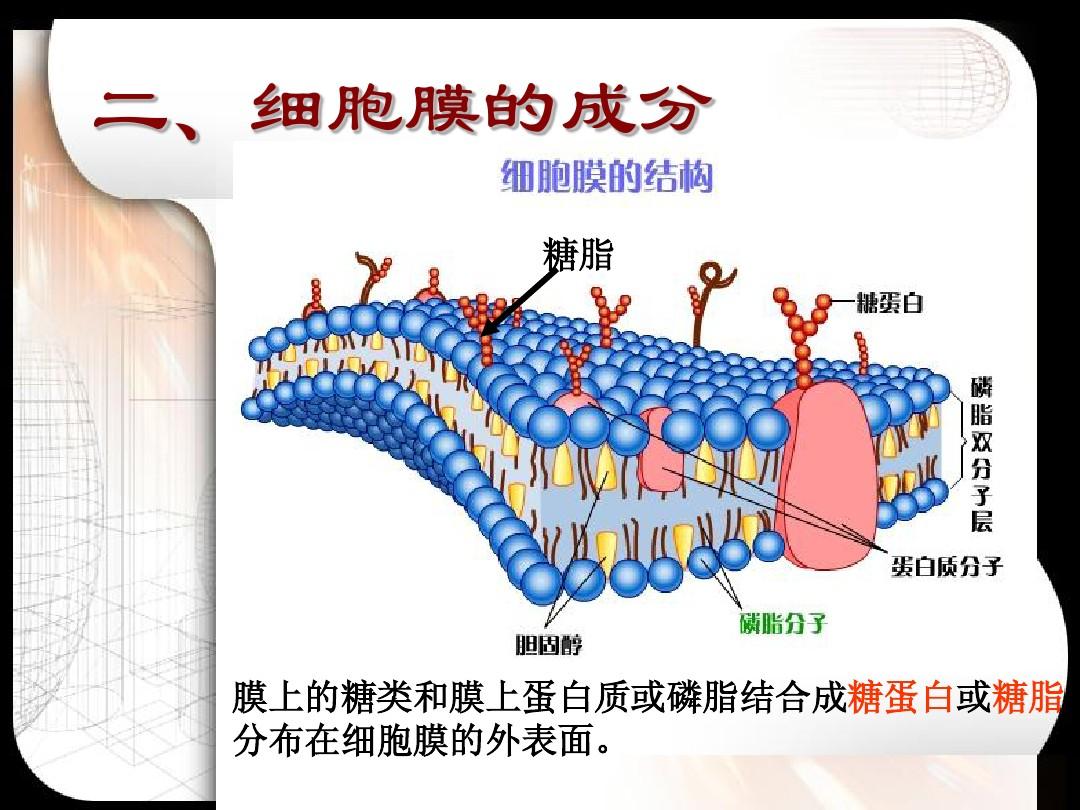
核纖層由核纖肽(lamin)構成,核纖肽分子量約60~80KD細胞膜是單層膜還是雙層膜,是一類中間纖維,在喂奶類和昆蟲中可分為A、B兩型。
作用
1.保持核的形態:是核被膜的支架,用高鹽氨水、非離子去污劑和核苷酸酶除去大部份核物質,剩余的核纖層仍能維持核的輪廓。據悉,核纖層與核骨架以及穿過核被膜的中間纖維相連,使胞質骨架和核骨架產生一連續網路結構。
2.參與染色質和核的組裝:核纖層在細胞分裂時呈現出周期性的變化,在間期核中,核纖層提供了染色質(異染色質)在核周邊錨定的位點。在前期結束時,核纖層被乙酸化,核膜解體。其中B型核纖肽與核膜殘余水泡結合,A型溶于胞質中。在分裂末期,核纖肽去乙酸化重新組裝,介導了核膜的重建。
二、核孔
核孔是半徑50~80nm的方形孔。內、外核膜在孔緣相連續,孔內有環()與中心顆粒組成核孔復合體。環有16個球狀亞單位,孔內、外線各有8個。從坐落核孔中心的中心顆粒(又稱孔栓)放射狀發出細絲與16個亞單位相連。核孔所在處無核纖層。通常覺得,水離子和核苷等小分子物質可直接通透核被膜;而RNA與蛋白質等大分子則經核孔出入核,但其出入方法尚不明了。其實,核功能活躍的細胞核孔數目多。成熟的卵子幾乎無核孔,而卵母細胞的核孔非常豐富,成為研究該結構的主要材料。
核孔是細胞核與細胞質之間物質交換的通道,一方面核的蛋白都是在細胞質中合成的,通過核孔定向輸入細胞核,另一方面細胞核中合成的各種RNA、核糖體亞單位須要通過核孔運到細胞質。據悉注射實驗證明,小分子物質才能以自由擴散的形式通過核孔步入細胞核。
核孔由起碼50種不同的蛋白質()構成,稱為核孔復合體(pore,NPC)。通常喂奶植物細胞平均有3000個核孔。細胞核活動旺盛的細胞中核孔數量較多,反之較少。如蛙卵細胞每位核可有37.7X106個核孔,但其成熟后細胞核僅150~300個核孔。
在電鏡下觀察,核孔是呈方形或八角形,通常覺得其結構如fish-trap,主要包括以下幾個部份:①胞質環(ring),坐落核孔復合體胞質兩側細胞膜是單層膜還是雙層膜,環上有8條纖維伸向胞質;②核質環(ring),坐落核孔復合體核質兩側,里面伸開8條纖維,纖維端部與端環相連,構成籠子狀的結構;③轉運器(),核孔中央的一個栓狀的中央顆粒;④輻(Spoke):核孔邊沿伸向核孔中央的突出物。
核被膜的物質運輸
1982年R.發覺核內含量豐富的核質蛋白()的C端有一個訊號序列,可引導蛋白質步入細胞核,也稱核定位訊號(,NLS)。第一個被確定的NLS是病毒SV40的T抗體,它在胞質中合成后很快積累在核中。其NLS為:pro-pro-lys-lys-lys-Arg-Lys-val,雖然單個多肽被替換,亦喪失作用。
NLS由4-8個多肽組成,富含Pro、Lys和Arg。對其聯接的蛋白質無特殊要求,但是完成核輸入后不被摘除。
是一類與核孔選擇性運輸有關的蛋白家族,相當于受體蛋白。其中負責將蛋白從細胞質運進細胞核,負責相反方向的運輸。
通過核孔復合體的轉運還涉及Ran蛋白,Ran是一種G蛋白,調節貨物受體復合體的組裝和解體,在細胞核內Ran-GTP的濃度遠低于細胞質。
核質蛋白向細胞核的輸入可描述如下:①蛋白與NLS受體,即α/β二聚體結合;②貨物與受體的復合物與NPC胞質環上的纖維結合;③纖維向核彎曲,轉運器構型發生改變,形結婚水通道,貨物通過;④貨物受體復合體與Ran-GTP結合,復合體解散,釋放出貨物;⑤與Ran-GTP結合的β,輸出細胞核,在細胞質中Ran結合的GTP酯化,Ran-GDP返回細胞核重新轉換為Ran-GTP;⑥α在核內的幫助下運回細胞質。
對細胞核向細胞質的大分子輸出了解較少,大多數情況下,細胞核內的RNA是與蛋白質產生RNP復合物轉運出細胞核的。RNP的蛋白質上具有核輸出訊號(,NES),可與細胞內的受體結合,產生RNP--Ran-GTP復合體,輸出細胞核后,Ran-GTP酯化,釋放出結合的RNA,Ran-GDP、和RNP蛋白返回細胞核。
核被膜染色觀察
在HE染色切塊上,細胞核()以其強嗜酸性而成為細胞內最顯眼的結構。因為它富含DNA--遺傳信息,因而,借DNA復制與選擇性轉錄,細胞核成為細胞增殖、分化、代謝等活動中關鍵環節之一。人體絕大多數種類的細胞具有單個細胞核,少數無核、雙核或多核。核的形態在細胞周期各階段不同,間期核的形態在不同細胞亦相差甚遠,但其結構都包括核被膜,染色質,胚乳與核基質四部。
核被膜觀察:在電鏡下觀察,可見核被膜由兩層平行排列的單位膜組成。內、外兩層膜之間的腔隙稱核周腔。核被膜內層表面有核蛋白體附著,并與核糖相接。核被膜上有許多由內、外膜層融合而成的孔,稱核孔。核孔是核內與細胞質之間進行物質交換的通道,核孔的數量可隨細胞生理狀況的不同而變化。核被膜對控制細胞質與核之間的物質交流,維持核內環境的恒定有重要作用。
核被膜核苷三乙酸酶()
核被膜上的活性與核膜結構的完整性息息相關,因而任何能影響細胞核膜結構的誘因幾乎都可以影響核膜活性。膳食中脂肪濃度可影響的活性與核膜中的脂類組成及尿酸濃度,喂食高P/S膳食的小鼠肝細胞核膜上C18:2ω6水平降低,酶活性下降;核膜上固醇及其氧化產物等均可直接影響的活性;JCR:LA-cp肥胖小鼠肝細胞核膜上尿酸濃度變化可調節其酶的活性。致畸物黃曲霉毒素、DM-NA等低劑量時可使酶活性降低,而致畸物高劑量時其毒性作用則可掩藏酶的活性變化,推斷酶活性變化可能是致畸劑使細胞核膜發生膨脹的結果,此可部份解釋個別癌癥發生過程中為何成熟mRNA要發生轉運的降低。同樣細胞核膜上自由基的變化也可影響核被膜的活性,從而影響成熟mRNA的轉運過程。
電鏡和螢光光子漂白掃描技術否認小鼠肝細胞核膜上存在胰島素結合位點,胰島素與其核膜上相應位點的結合后,可通過核膜上酶的活性變化,和核孔樣收縮器的定向作用來影響生物大分子包括mRNA在內的特異性出核轉運;同樣核膜上有乳腺素和甾體結合位點以及EGF受體,它們在與其官能團結合后,通過核膜空間結構的改變或則通過其它途徑,影響活性和生物分子的跨核膜轉運。6月齡肥胖雄性小鼠肝細胞核膜酶活性顯著低于相應小鼠NT-Pase的活性,且的Vmax值隨年紀的降低而降低,提示其活性變化可能是體內雌激素經核膜受體作用的結果。
分子生物學的發展尤其是雜交技術的簡化使人們能從mRNA水平推算蛋白質的抒發情況,而且mRNA和蛋白質的變化發展似乎是不平行的。自從細胞核膜被發覺與確認以來,人們越來越倍感要研究蛋白功能的實驗必須補充了解mRNA的特異轉運。Gupta報導小鼠心肌細胞核膜受去乙酸化作用調節,因而由激素或其它因子造成訊號轉導作用,經細胞核膜的活性變化調節mRNA的特異轉運,可能會成為今后進一步研究代謝調控的重點。
伴護分子p97對核被膜的改建
伴護分子p97是一種ATP酶,參與各類不同的功能,如核糖中蛋白的降解和細胞周期調控等。在有絲分裂開始的時侯,核被膜被拆開,并在過程的最后被整修。這項研究為核被膜的整修提供了一個機理上的解釋。研究發覺,p97與染色質上B(核被膜產生的一種已知的抑制成份)的一種泛素化方式相結合。這造成B從染色質中被提取下來,因而容許染色體解凝和核被膜產生。