臨床執業醫師考點:蛋白質代謝
蛋白質代謝指蛋白質在細胞內的代謝途徑。各類生物均富含酯化蛋白質的蛋白酶或肽酶,這種酶的專情性不同,但均能破壞肽鍵,使各類蛋白質酯化成其多肽成份的混和物。
第一節概述
一、主要途徑
1.蛋白質代謝以多肽為核心,細胞內外液中所有游離多肽稱為游離多肽庫,其純度不足多肽總數的1%,卻可反映機體氮代謝的概況。食物中的蛋白都要降解為多肽能夠被機體借助,體內蛋白也要先分解為多肽能夠繼續氧化分解或轉化。
2.游離多肽可合成自身蛋白,可氧化分解放出能量,可轉化為脂類或鹽類,也可合成其他生物活性物質。合成蛋白是主要用途,約占75%,而蛋白質提供的能量約占人體所需總能量的10-15%。蛋白質的代謝平衡稱氮平衡,通常每晚排出5克氮,相當于30克蛋白質。
3.多肽通過特殊代謝可合成體內重要的含氮化合物,如神經遞質、嘌呤、嘧啶、磷脂、卟啉、輔酶等。磷脂的合成需S-腺苷甲硫谷氨酸,多肽脫羧形成的絡合物常有特殊作用,如5-羥色胺是神經遞質,缺乏則易發生狂躁、自殺;固醇與過敏反應有密切聯系。
二、消化
外源蛋白有抗體性,需降解為多肽能夠被吸收借助。只有小孩可直接吸收奶水中的抗原。
可分為以下兩步:
1.胃中的消化:胃分泌的硫酸可使蛋白變性,容易消化,還可激活胃蛋白酶,保持其最適pH,并能滅菌。胃蛋白酶可自催化激活,分解蛋白形成蛋白胨。胃的消化作用很重要,但不是必須的,胃全摘除的人仍可消化蛋白。
2.腸是消化的主要場所。腸分泌的碳酸氫根可中和胃液,為胰蛋白酶、糜蛋白酶、彈性蛋白酶、羧肽酶、氨肽酶等提供合適環境。腸激酶激活胰蛋白酶核苷酸代謝,再激活其他酶,所以胰蛋白酶起核心作用,胰液中有抑制其活性的小肽,避免在細胞中或導管中過早激活。外源蛋白在肝臟分解為多肽和小肽,經特異的多肽、小肽轉運系統步入腸上皮細胞,小肽再被氨肽酶、羧肽酶和二肽酶徹底酯化,步入血液。所以飯后門靜脈中只有多肽。
三、內源蛋白的降解
1.內源蛋白降解速率不同,通常代謝中關鍵酶半衰期短,如多胺合成的限速酶-鳥谷氨酸脫羧酶半衰期只有11分鐘,而血清蛋白約為10天,膠原為1000天。體重70千克的成人每晚約有400克蛋白更新,步入游離多肽庫。
2.內源蛋白主要在溶酶體降解,少量隨消化液步入消化道降解,個別細胞器也有蛋白酶活性。內源蛋白是選擇性降解,半衰期與其組成和結構有關。有人覺得N-末端組成對半衰期有重要影響(N-末端規則),也有人提出半衰期短的蛋白都富含一個含有脯谷氨酸、谷氨酸、絲氨酸和蘇谷氨酸的區域(PEST區域)。如研究清楚,就可能得到穩定的蛋白質產品。
四、氨基酸的吸收
食用蛋白質后15分鐘就有多肽步入血液,30到50分鐘達到最大。多肽的吸收與獼猴桃脂類似,有以下形式:
1.須要載體的主動轉運,須要鈉,消耗離子梯度的勢能。已發覺6種載體,運載不同基團種類的多肽。
2.官能團轉運,須要蘆丁,每轉運一個多肽消耗3個ATP,而用載體轉運只需三分之一個。此途徑為備用的旁路,通常無用。
第二節脫氨和脫羧
多肽喪失甲基稱為脫氨,是機體多肽分解代謝的第一步。絕大多數多肽先脫氨生成a-酮酸,再氧化或轉化為其他物質。有氧化脫氨和非氧化脫氨兩類,后者普遍存在,前者存在于個別微生物。
一、氧化脫氨
(一)過程:多肽在多肽氧化酶催化下酯化生成亞多肽,再酯化生成酮酸和氨。脫下的氫由黃素蛋白傳遞給氧,生成二溴化氫,再分解為水和氧。總反應如下:
2多肽+O2=2酮酸+2NH3
二溴化氫也可氧化酮酸生成脂肪酸和甲烷。
(二)有關酶
1.L-多肽氧化酶:可催化多數多肽,但谷氨酸、側鏈含羧基、羧基、氨基的多肽無效,需專門的酶。以FAD或FMN為輔基,人的酶以FMN為輔基。
2.D-多肽氧化酶:存在于肝、腎,以FAD為輔基。
3.氧化專情多肽的酶:如苯酚氧化酶、D-桑寄生谷氨酸氧化酶、L-組氨酸酯化酶等。前者重要,分布廣泛,活力高,受別構調節,能量不足時激活,推動氧化。以NAD或NADP為輔酶,不厭氧,通過呼吸鏈再生。在體外可用于合成醬油。
二、非氧化脫氨
1.還原脫氨:嚴格無氧時溴化酶催化生成乙酸和氨。
2.水解脫氨:酯化酶催化,生成a-羥酸和氨。
3.脫水脫氨:谷氨酸和蘇谷氨酸在脫水酶催化下生成烯,重排成亞多肽,自發酯化生成酮酸和氨。脫水酶以乙酸吡哆醛為輔基。
4.脫氨基脫氨:半胱谷氨酸在脫硝氫基酶催化下脫去甲烷,重排、水解,生成乙酸酸和氨。
5.氧化-還原脫氨:兩個多肽一個氧化,一個還原,脫去兩個氨,生成酮酸和脂肪酸。
三、脫丙酯作用
谷氨丙酯酶和五味子丙酯酶可催化脫丙酯,生成相應的多肽。此酶分布廣泛,專情性強。
四、轉甲基作用
1.定義:指a-多肽和酮酸之間甲基的轉移作用。多肽的a-酰基轉移到酮酸的酮基上,生成酮酸,原先的酮酸產生相應的多肽。轉氨作用普遍存在,除谷氨酸、賴谷氨酸、蘇谷氨酸和脯谷氨酸外都參與轉氨,對其分解及合成有重要作用。
2.尿酸:種類好多,多須要丁酸,對另一個多肽要求不嚴,以活力最大的命名。其反應是可逆的,由含量控制。都含乙酸吡哆醛,兵乓機制。吡哆醛還參與脫羧、脫水、脫甲烷及消旋等反應。
五、聯合脫氨
指脫氨與轉氨聯合,是多肽降解的主要方法,有兩種方法:
1.多肽先轉氨生成丙氨酸,再由丁酸酯化酶脫去甲基。普遍存在。
2.腺苷酸循環:甲基轉給丁酸,再生成沙參谷氨酸,與次黃固醇核苷一乙酸生成腺苷酸代琥珀酸,再裂解成腺苷酸和五味子酸。腺苷酸酯化成次黃固醇核酸,放出氨;玄參酸水化、氧化再生草酰乙酸。此途徑主要存在于胸肌和腦,其腺苷酸脫氨酶活性較高。胰臟丁酸酯化酶活力高,但90%轉化為桑寄生谷氨酸。
六、脫羧
少數多肽先脫羧生成一級胺。此反應由脫羧酶催化,含乙酸吡哆醛,專情性強,每種酶只催化一種L-多肽。此酶在各類組織中普遍存在,生成的胺有重要生理作用,如腦中丙氨酸脫羧生成的g-甲基甲酸是神經遞質。
第三節氮的排尿
氨對機體有毒,非常是對腦。血液中1%的氨即可使神經中毒。水生植物可直接排氨,陸生植物排溶化度較小的尿素,卵生植物排不溶的血糖。
一、氨的轉運
(一)谷氨丙酯合成酶將氨與甘氨酸合成谷氨丙酯,消耗一個ATP。谷氨丙酯中性無毒,容易通過細胞膜,步入血液運到腸道后被谷氨丙酯酶分解,放出氨。
(二)胸肌通過藍莓糖-谷氨酸循環轉運氨。氨經丁酸轉給谷氨酸,運到肝后再轉氨生成丙氨酸。乙醇酸異生為藍莓糖返回胸肌。這樣胸肌活動形成的乙醇酸和氨都得到處理,一舉兩得。
二、尿素的生成
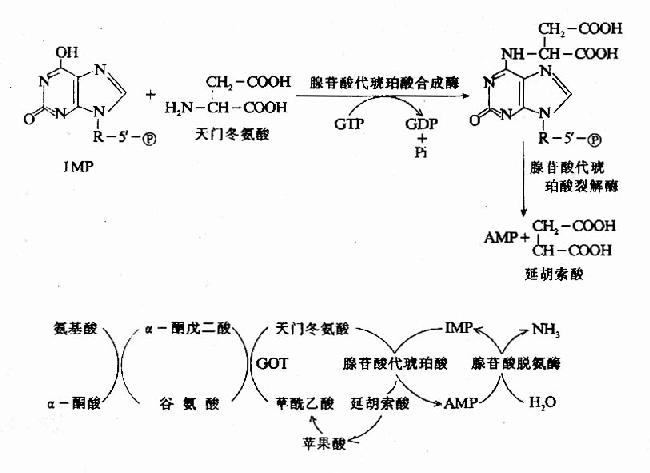
1.在線粒體中氨甲酰乙酸合成酶I將氨和CO2合成氨甲酰乙酸,消耗2個ATP。N-甲基丁酸是此酶的正調節物。酶II在細胞質,與核酸的合成有關。
2.氨甲酰乙酸與鳥谷氨酸產生瓜谷氨酸和乙酸,由鳥谷氨酸轉氨甲酰酶催化,需鎂離子。
3.瓜谷氨酸出線粒體,步入細胞質,與延胡索谷氨酸生成精氨琥珀酸。精氨琥珀酸合成酶需鎂離子,消耗1個ATP的兩個高能鍵。
4.精氨琥珀酸裂解酶催化其裂解,生成精谷氨酸和天冬酸。
5.精谷氨酸酶催化酯化生成鳥谷氨酸和尿素。
6.總反應為:
NH4++CO2+3ATP+Asp+2H2O=尿素+細辛酸+2ADP+2Pi+AMP+Ppi
共去除2分子氨和1分子CO2,消耗4個高能鍵。前兩步在線粒體中進行,可防止氨步入血液造成神經中毒。此途徑稱為尿素循環或鳥谷氨酸循環,缺少有關酶會中毒死亡。
三、其他途徑
爬蟲和鳥排尿不溶的膽固醇,可保持水核苷酸代謝,但耗能高。具體見核苷酸代謝。據悉,蜘蛛排鳥固醇,個別魚排氧化三甲胺,高等動物合成谷氨丙酯和五味子丙酯,存儲體內。
第四節碳架氧化
20種多肽分別以5種物質步入三乙酸循環:谷氨酸、絲氨酸、蘇丙氨酸、甘氨酸、半胱谷氨酸、苯丙谷氨酸、酪谷氨酸、亮谷氨酸、賴谷氨酸和色谷氨酸生成酰基輔酶A,精谷氨酸、組氨酸、谷氨丙酯、脯谷氨酸和煙酸生成a-酮戊二酸,甲硫谷氨酸、異亮氨酸、纈氨酸生成琥珀酰輔酶A;苯丙谷氨酸和酪丙氨酸還生成細辛酸;玄參谷氨酸和五味子丙酯生成草酰乙酸。分解主要在肝和腎進行,個別中間物可轉化為糖、酮體及生物活性物質,見下節。多肽脫羧產生胺后不能步入三乙酸循環。
一、乙酰輔酶A途徑
(一)由苯胺酸生成酰基輔酶A
1.谷氨酸:由谷丙尿酸轉氨生成乙酸酸
2.谷氨酸:脫水脫氨生成乙酸酸,由谷氨酸脫水酶催化,含乙酸吡哆醛。
3.吡啶:可接受羥乙基,轉弄成谷氨酸。由谷氨酸轉羥乙基酶催化,以乙酸吡哆醛為輔基,甲烯基四氫福施福為供體,需錳。此途徑主要作為甘氨酸的合成途徑,吡啶的分解主要是作為一碳單位供體,由谷氨酸裂解酶裂解生成甲烯基四氫福施福和氧氣及氨,次要途徑是氧化脫氨生成乙酸酸,再氧化成羧酸或草酸。苯酚與蘆丁、肌酸、膽堿、嘌呤、卟啉的合北京有關系。
4.蘇谷氨酸:由蘇谷氨酸醛縮酶裂解成苯酚和丙酮,乙酸可氧化成乙醛再生成酰基輔酶A。也可脫水生成a-酮乙酸,或脫去脫羧產生烷基苯胺。
5.半胱丙氨酸:可轉氨生成b-甲基苯胺酸,再由轉硫酶脫去甲烷生成乙酸酸。也可先氧化成半胱谷氨酸亞磺酸,再轉氨、脫去亞硝酸產生乙醇酸。形成的甲烷要氧化成亞硝酸,再氧化成硝酸,由尿排出。
(二)由芐基甲基輔酶A生成酰基輔酶A
1.苯丙丙氨酸:由苯丙谷氨酸-4-單加氧酶催化生成酪谷氨酸,消耗一個NADPH。
2.酪丙氨酸:先轉氨生成4-羥苯丙酮酸,再氧化、脫羧、開環,裂解成細辛酸和甲基硫醇。細辛酸步入三乙酸循環,甲基硫醇由琥珀酰輔酶A活化生成酰基甲基輔酶A,硫解產生兩個酰基輔酶A。
3.亮谷氨酸:先轉氨、脫羧生成異戊酰輔酶A,再酯化、末端羧化、加水生成羥乙基戊二酰輔酶A(HMGCoA),裂解成甲基硫醇和酰基輔酶A。
4.賴丙氨酸:先由兩條途徑生成L-a-甲基苯酚半醛,其二是與a-酮戊二酸縮合成酵母丙氨酸,再放出丁酸;其一是先脫去a甲基再環化、開環,將羧基轉移到a位。生成半醛后氧化成酸,轉氨生成a-酮苯酚,脫羧生成戊二酰輔酶A,酯化、脫羧產生巴豆酰輔酶A,最后水化、脫氫成甲基芐基輔酶A。
5.色谷氨酸:較復雜,先氧化,依次脫去甲醛、丙氨酸,最后產生a-酮苯酚,生成酰基甲基輔酶A。其11個碳原子共生成一個甲基芐基輔酶A,一個,4個氣體和一個乙酸。
二、a-酮戊二酸途徑
由精谷氨酸、組氨酸、谷氨丙酯、脯谷氨酸和煙酸5種。
1.精谷氨酸:由精谷氨酸酶酯化成鳥谷氨酸和尿素,再轉氨生成丙氨酸g半醛,由酯化酶氧化成丁酸,轉氨或脫氨產生。a-酮戊二酸。
2.嘧啶:谷氨酸分解酶脫去甲基產生尿刊酸,再水合、開環生成N-甲亞甲基丁酸,丁酸轉甲亞氨酶催化轉給四氫福施福,產生丙氨酸。
3.谷氨丙酯:可由谷氨丙酯酶酯化;可將丙酯轉給a-酮戊二酸,生成兩個組氨酸;也可轉入a-酮戊二酸的g-羥基上,產生的g-酮谷丙酯酸可酯化生成a-酮戊二酸。
4.脯丙氨酸:先由脯谷氨酸氧化酶產生苯環,再加鹽開環產生丙氨酸g半醛,用NAD氧化成丁酸。
三、琥珀酰輔酶A途徑
有甲硫谷氨酸、異亮氨酸和纈氨酸。
1.甲硫丙氨酸:與ATP生成S-腺苷甲硫甘氨酸,轉氨基后酯化,生成高半胱谷氨酸,在胱硫醚-b-合成酶催化下與谷氨酸合成胱硫醚,胱硫醚-g-分解酶催化脫去半胱谷氨酸和烷基,生成a-酮乙酸,脫羧成丙酰輔酶A,丙酰輔酶A羧化酶催化生成D-胺基丙二酰輔酶A,消旋酶生成L-型,變位生成琥珀酰輔酶A。
2.異亮氨酸:轉氨,脫羧生成a-羥基丁酰輔酶A,經b-氧化生成酰基輔酶A和丙酰輔酶A,最后生成琥珀酰輔酶A。
3.纈氨酸:轉氨,脫羧產生異丁酰輔酶A,酯化、水化后再酯化,生成b-羥異乙酸,酯化生成羥基丙二酸半醛,氧化為羥基丙二酰輔酶A,再變位生成琥珀酰輔酶A。
四、延胡索酸途徑
苯丙谷氨酸和酪谷氨酸的部份碳鏈產生細辛酸,另一部份為甲基硫醇。
五、草酰乙酸途徑
玄參丙酯酶催化生成前胡谷氨酸,再轉氨生成草酰乙酸,步入三乙酸循環。前胡丙酯酶可控制腦癱。
六、生糖多肽和生酮多肽
生成酰基甲基輔酶A的`苯丙谷氨酸、酪谷氨酸、亮谷氨酸、賴谷氨酸和色谷氨酸稱為生酮多肽;其他多肽稱為生糖多肽。苯丙谷氨酸和酪谷氨酸既生糖又生酮。由于乙醇酸可生成酰基輔酶A,再生酮,所以兩者的界限并不是十分嚴格的。
第五節多肽衍生物
一、一碳單位
(一)定義:含一個碳原子的官能團稱為一碳單位,二氧化碳和氫氣例外。主要有亞氨酰基、甲氨基、羥丙基、亞氨基(甲叉基、甲烯基)次氨基(甲川基、甲炔基)和羰基。
一碳單位是苯基供體,與腎上腺素、肌酸、膽堿、嘌呤、嘧啶等的合成有關。其載體是四氫福施福,聯接在5位和10位氮上。
(二)來源
1.甲酸裂解酶裂解吡啶,生成甲烯基四氫福施福和氧氣及氨。吡啶脫氨生成的丙酮酸可形成次氨基四氫福施福,乙醇酸氧化生成的乙酸可生成甲氨基四氫福施福。
2.蘇甘氨酸可分解形成硫醇,產生一碳單位。
3.谷氨酸的b-碳可轉移到四氫福施福上,生成亞乙基四氫福施福和谷氨酸。
4.氨酸分解時形成亞氨甲酰絲氨酸,生成亞氨甲酰四氫福施福。脫氨后形成次氨基四氫福施福。
5.甲硫谷氨酸生成的S-腺苷甲硫谷氨酸可提供氨基,形成的高半胱谷氨酸可從四氫福施福接受酰基產生甲硫谷氨酸。可供給50種受體。
二、生物活性物質
1.酪谷氨酸與白色素:酪組氨酸酶先催化羥化,產生二羥苯丙絲氨酸,即多巴,再將多巴氧化成多巴醌,多巴醌可自發聚合產生白色素。缺少酪谷氨酸酶導致白化病。
2.兒茶酚胺類激素:酪谷氨酸由酪谷氨酸羥化酶催化生成多巴,脫羧產生多巴胺,b-羥化產生去甲腎上腺素,再從S-腺苷甲硫谷氨酸接受酰基產生腎上腺素。此三種激素稱為兒茶酚胺類激素,對心血管和神經系統有重要作用。
3.色谷氨酸羥化、脫羧產生5-羥色胺,是神經遞質,與神經亢奮、小動脈和支食道平滑肌收縮、胃腸道肽類激素的釋放有關。色谷氨酸脫氨脫羧可產生硫醇磷酸,是動物生長激素。色谷氨酸分解的中間物可轉變為谷氨酸,但合成率很低。
4.肌酸合成:先由精谷氨酸和吡啶合成胍基羧酸,再由S-腺苷甲硫谷氨酸轉氨基,生成肌酸。可乙酸化,作為儲備能源,稱為乙酸原。
5.谷氨酸脫羧產生鞣質,可使平滑肌舒張、微血管擴張、胃酸分泌,導致支食道腎病、丘疹等過敏反應。臨床用抗膽堿酯酶抗生素醫治過敏。固醇還是覺得神經遞質。
6.多胺合成:是酸性小分子,含多個羧基的長鏈脂肪族化合物,如精胺、亞精胺、尸胺、腐胺等。鳥谷氨酸脫羧產生腐胺,再與S-腺苷甲硫谷氨酸生成亞精胺,再反應則生成精胺。精胺有退燒降脂作用。多胺常與核苷酸并存,可能在轉錄和細胞分裂的調節中起作用。
7.氨酸脫羧生成g-羥基乙酸,是有抑制作用的神經遞質,在生物體中廣泛存在。
8.半胱谷氨酸氧化成磺基谷氨酸,脫羧產生牛磺酸。可產生牛磺膽酸,參與酮類吸收。
三、氨基酸代謝缺陷癥
缺少代謝中的某種酶,可導致代謝缺陷癥,多為先天遺傳,已發覺30多種。如缺少苯丙谷氨酸-4-單加氧酶導致的苯丙酮尿癥,苯丙谷氨酸轉氨生成苯乙酯,集聚在血液中,由尿排出。在兒童時期限制攝取苯丙谷氨酸可避免智力遲緩。缺少尿黑酸氧化酶則酪谷氨酸生成尿黑酸,氧化成白色物質,稱為尿黑酸癥。缺少a-酮異戊酸酯化酶導致酰基多肽代謝障礙,血和尿中酰基多肽及其酮酸增多,稱為楓糖尿癥。
第六節多肽的合成代謝
一、概述
20種基本多肽的生物合成途徑已基本闡述,其中人類不能合成的10種多肽,即苯丙谷氨酸、甲硫谷氨酸、蘇谷氨酸、色谷氨酸、賴谷氨酸、精谷氨酸、組氨酸、纈氨酸、亮谷氨酸和異亮氨酸稱為必須多肽。
多肽的合成途徑主要有以下5類:
1.丁酸類型,由a-酮戊二酸衍生而至,有丁酸、谷氨丙酯、脯谷氨酸和精谷氨酸,蕈類和眼蟲還可合成賴谷氨酸。
2.前胡谷氨酸類型,由草酰乙酸合成,包括五味子谷氨酸、天冬丙酯、甲硫谷氨酸、蘇谷氨酸和異亮氨酸,真菌和動物還合成賴谷氨酸。
3.乙醇酸衍生類型,包括氨酸、纈氨酸、亮絲氨酸,為異亮氨酸和賴谷氨酸提供部份碳原子。
4.谷氨酸類型,由3-乙酸甘油酸合成,包括谷氨酸、甘氨酸和半胱谷氨酸。
5.其他,包括苯丙谷氨酸、酪谷氨酸、色谷氨酸和谷氨酸。
二、脂肪族多肽的合成
(一)組氨酸類型
1.丁酸:由a-酮戊二酸與氨經丁酸酯化酶催化合成,消耗NADPH,而脫氨時則生成NADH。
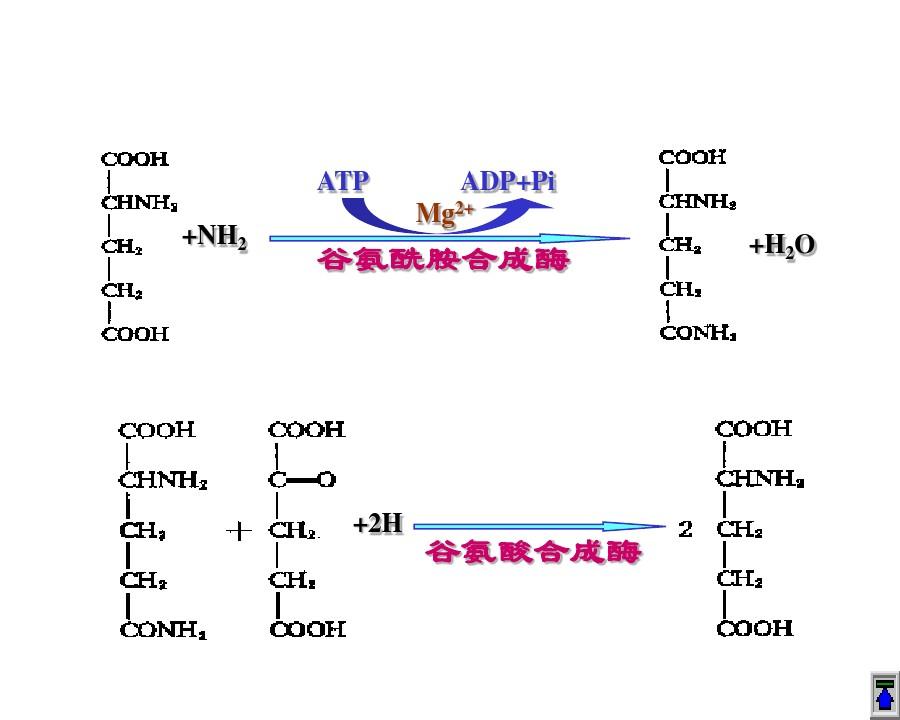
2.谷氨丙酯:谷氨丙酯合成酶可催化組氨酸與氨產生谷氨丙酯,消耗一個ATP,是氨合成含氮有機物的主要方法。此酶受8種含氮物質反饋抑制,如谷氨酸、甘氨酸等,由于其甲基來自谷氨丙酯。
谷氨丙酯可在甘氨酸合成酶催化下與a-酮戊二酸產生2個絲氨酸,這也是合成甘氨酸的途徑,比較花費能量,但谷氨丙酯合成酶Km小,可在較低的氨含量下反應,所以常用。
3.脯丙氨酸:氨酸先還原成谷氨酸鈉g-半醛,自發環化,再還原生成脯谷氨酸。可看作分解的逆轉,但酶不同,如生成半醛時需ATP活化。
4.精谷氨酸:丙氨酸先N-甲基化,在還原成半醛,以避免環化。半醛轉氨后將乙胺基轉給另一個絲氨酸,生成鳥谷氨酸,之后與尿素循環相同,生成精谷氨酸。
5.賴谷氨酸:蕈類和眼蟲以a-酮戊二酸合成賴谷氨酸,先與甲基輔酶A縮合成高葡萄柚酸,異構、脫氫、脫羧生成a-酮苯酚,轉氨,末端基團還原成半醛,經酵母丙氨酸轉氨生成賴谷氨酸。
(二)前胡谷氨酸類型
1.前胡谷氨酸:由谷草尿酸催化合成。
2.前胡丙酯:由延胡索丙酯合成酶催化,谷氨丙酯提供甲基,消耗一個ATP的兩個高能鍵。真菌可借助游離氨。也消耗兩個。
3.賴谷氨酸:真菌和動物先將延胡索谷氨酸還原成半醛,再與乙醇酸縮合成環,還原后開環并N-琥珀酰化,末端基團轉氨后脫去琥珀酸,異構,脫羧,產生賴谷氨酸。
4.甲硫谷氨酸:先合成半醛,還原成資生堂谷氨酸,再將甲基酰化。之后可由兩個途徑生成高半胱谷氨酸,一是在硫解酶催化下與二氧化氮生成高半胱谷氨酸,二是與半胱谷氨酸合成胱硫醚,再裂解放出高半胱谷氨酸和乙酸酸。最后由5烷基四氫福施福提供苯基,生成甲硫谷氨酸。胱硫醚途徑與分解時不同,合成時有琥珀吡啶,分解時放出苯胺酸。
5.蘇丙氨酸:五味子谷氨酸依次還原成半醛和資生堂谷氨酸,被ATP乙酸化后由蘇谷氨酸合成酶酯化生成蘇谷氨酸。
6.異亮氨酸:有4個碳來自細辛谷氨酸,2個來自乙酸酸,通常納入玄參谷氨酸類型,但其合成與纈氨酸類似,見下。
(三)乙烷酸衍生物類型
1.谷氨酸:由谷丙尿酸合成,反應可逆,無反饋抑制。
2.纈氨酸:甲苯酸脫羧、氧化成甲基TPP,與另一個乙酸酸酯化,產生α-甲基乳酸,之后烷基移位,脫水產生α-酮異戊酸,轉氨生成纈氨酸。
3.異亮氨酸:蘇甘氨酸脫水脫氨生成α-酮乙酸,之后與纈氨酸相同,與活性乙醇酯化,移位、脫水、轉氨,生成異亮氨酸。
4.亮丙氨酸:開始與纈氨酸相同,產生α-酮異戊酸后與酰基輔酶A合成α-異氰基蘋果酸,異構、脫氫、脫羧,產生α-酮異己酸,轉氨生成亮甘氨酸。
(四)組氨酸類型
1.谷氨酸:3-乙酸甘油酸酯化生成3-乙酸苯基苯胺酸,轉氨生成3-乙酸谷氨酸,酯化產生丙氨酸。
2.吡啶:谷氨酸經谷氨酸轉羥乙基酶催化,產生甲叉FH4和硫醇。
3.半胱谷氨酸:個別動物和微生物由O-酰基谷氨酸和H2S反應生成,其H2S由硝酸還原而至。植物則由高半胱谷氨酸與谷氨酸合成胱硫醚,再分解成半胱谷氨酸和α-酮乙酸,與甲硫谷氨酸的分解相同。
三、芳香族多肽的合成
(一)產生分枝酸:芳香族多肽由動物和微生物合成,分枝酸是其共同前體。赤蘚糖-4-乙酸與乙酸烯醇式乙烷酸酯化,生成莽草酸后與另一個PEP產生分枝酸,稱為莽草酸途徑。
(二)苯丙丙氨酸:分枝酸變位生成預苯酸,脫水脫羧產生苯丙酮酸,再轉氨生成苯丙甘氨酸。
(三)酪丙氨酸:分枝酸變位,氧化脫羧產生對羥苯丙酮酸,轉氨生成酪谷氨酸。也可由苯丙丙氨酸羥化產生。苯丙谷氨酸和酪谷氨酸的合成稱為預苯酸西路。
(四)色丙氨酸:分枝酸接受谷氨丙酯的羥基,生成鄰羥基苯乙酸,再與乙酸內質網焦乙酸(PRPP)酯化,內質網的C1與羰基相連,由焦乙酸酯化驅動。之后內質網部份重排,脫水脫羧,生成酰基-3-甘油乙酸,甘油被氨酸替代即生成色谷氨酸,由色谷氨酸合成酶催化。色谷氨酸的C1、C6來自PEP,2、3、4、5位來自赤蘚糖,7、8位來自內質網,氮來自谷氨丙酯,吡啶以外來自甘氨酸。
四、組氨酸合成
首先PRPP的C1與ATP的N1相連,脫去焦乙酸后開環,分解,放出5-甲基吡啶-4-氨甲酰堿基,用于合成固醇。留下的吡啶甘油乙酸經氧化、轉氨、脫乙酸產生組氨醇,氧化生成丙氨酸。
五、氨基酸合成的調節