撰文|LB
2021年1月25日,發表了美國哥廷根學院題為ofover500yearsofplant的研究論文。該文章研究了一個鞘脂脂肪酸不飽和酶編碼基因PpSFD介導小立碗蘚耐冷性的機制,并與擬南芥做了比較,闡明了苔蘚與種子動物調控細胞膜流動性的趨同進化機制。
過去二六年,苔蘚動物小立碗蘚(())早已成為研究動物進化發育生物學、植物分子生物學、植物比較基因組學等領域的模型系統。為何小立碗蘚會得到動物科學家們的追捧呢?主要有以下幾個方面的緣由:(1)苔蘚動物具有相對簡單的發育模式,其單倍體的胚珠體在生活史中占主導地位,這促使在研究其基因功能方面具有奇特的優勢;(2)二六年前,人們發覺苔蘚動物小立碗蘚就能高效地通過同源重組的方法將外源DNA整合到其核基因組中,這促使在小立碗蘚中舉辦反向遺傳學實驗顯得簡單易行;(3)小立碗蘚是第一個進行基因組測序的非種子動物,加上越來越多的分子遺傳學工具成功應用到該研究系統細胞膜流動性,促進了這一模型系統的應用;(4)小立碗蘚具有很強的抗旱、抗冷和抗鹽脅迫的能力細胞膜流動性,也是逆境生物學研究的理想材料。
Fig.4:ofP.type
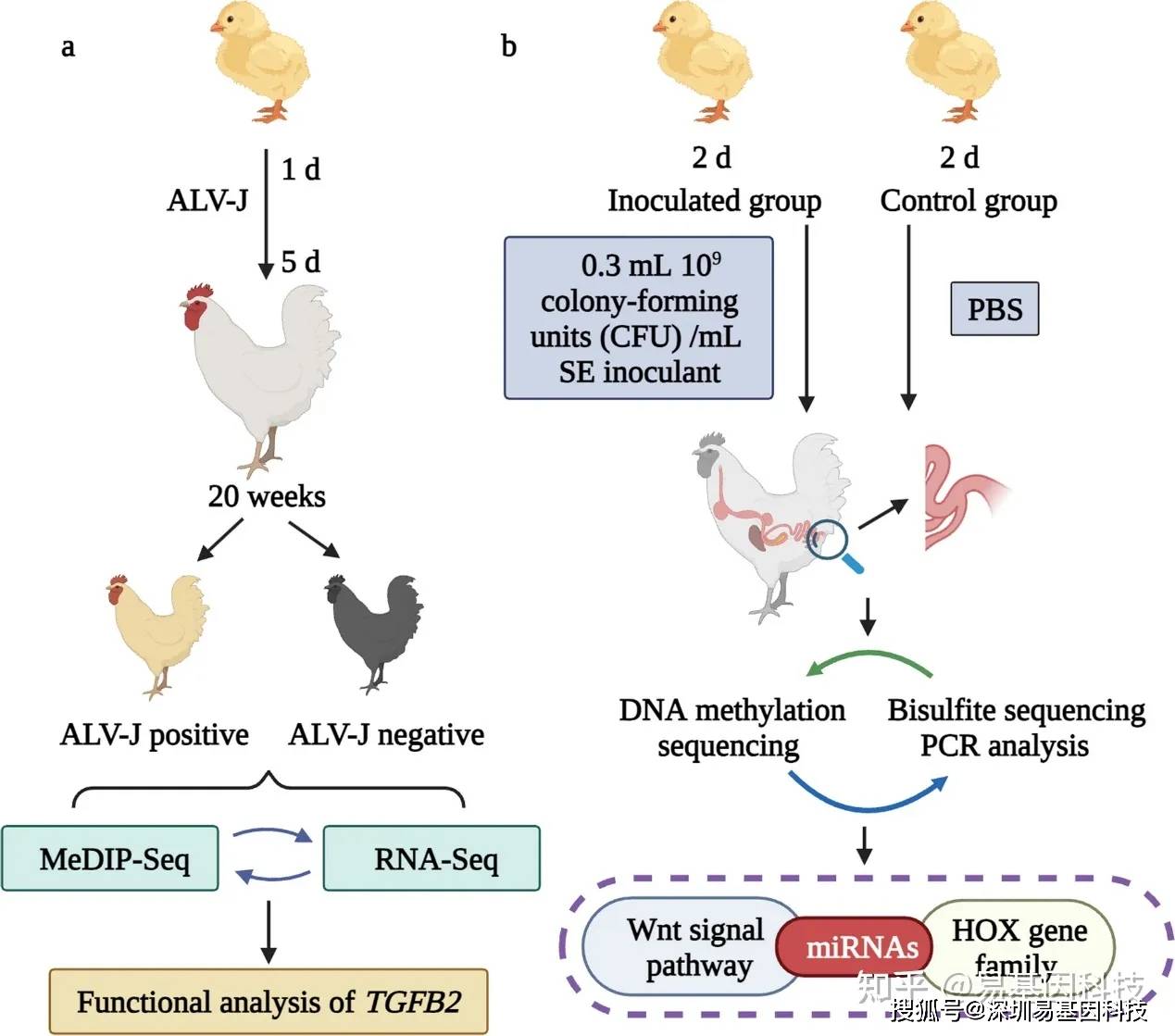
在本論文中,作者以小立碗蘚為系統,研究了一個鞘脂脂肪酸不飽和酶編碼基因PpSFD介導小立碗蘚耐冷性產生的分子機制。作者發覺在PpSFD基因無功能突變體的脂類譜中缺少富含單不飽和C24脂肪酸的鞘脂,表現為對冷敏感和對卵菌腐霉易感,表明該基因可能通過修飾磷脂從而影響細胞膜的流動性、參與了小立碗蘚的脅迫耐受能力產生。
Fig.5:Cold-withA.
有趣的是,該基因在擬南芥.1(甲基輔酶A去飽和酶類基因)突變體中異位抒發才能在功能上恢復該突變體的冷敏感表型,顯示它們具有類似的生物學功能即催化一個類似的反應。并且,這兩個酶具有顯著不同的進化起源,是一個氨基末端去飽和酶,而PpSFD是細胞色素b5融合去飽和酶。
Fig.7:AfortheofPpSFD
綜上,作者覺得在苔蘚與種子動物這兩個分化于5億年前的動物類群中,對細胞膜流動性的調控是獨立進化的,這是一個典型的趨同進化案例。
原文鏈接:
本推文未經專家審讀,限于作者水平,難免存在錯漏,僅供參考!如有疑惑,請閱讀原文獻!