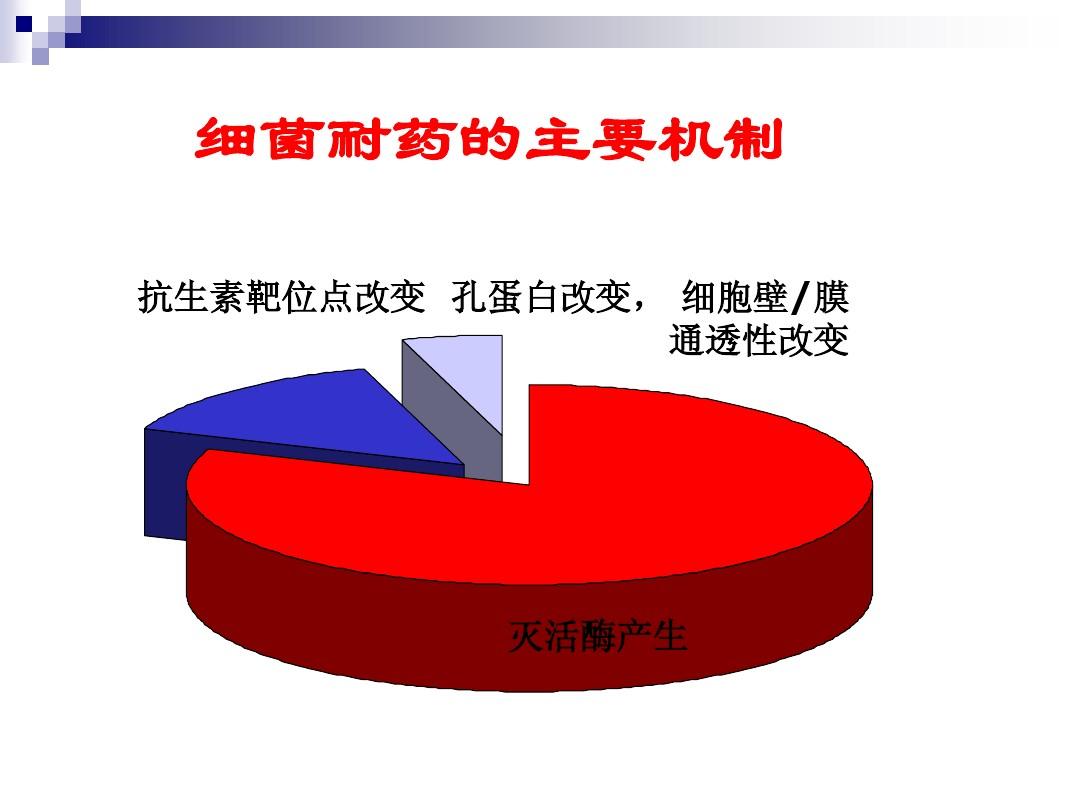
微生物檢驗必須把握的三大耐藥機制
你曉得哪些是微生物檢驗嗎?你對微生物檢驗了解嗎?下邊是我為你們帶來的關于微生物檢驗必需要曉得的三大耐藥機制的知識,歡迎閱讀。
一、產生滅活藥物的各類酶
1、β—內丙酯酶(β-)
β—內丙酯類藥物都共同具有一個核心β—內丙酯環,其基本作用機制是與真菌的抗生素結合蛋白結合,進而抑制真菌細胞壁的合成。形成β—內丙酯酶是真菌對β-內丙酯類抑菌抗生素形成耐藥的主要誘因。真菌形成的β-內丙酯酶,可利用其分子中的谷氨酸活性位點,與β—內丙酯環結合并打開β—內丙酯環,造成抗生素失活。迄今為止報導的β—內丙酯酶已超過300種,1995年Bush等將其分為四型:第1型為不被克拉維酸抑制的阿莫西林菌素酶;第2型為能被克拉維酸抑制的β-內丙酯酶;第3型為不被所有β—內丙酯酶抑制劑抑制的金屬β-內丙酯酶(需Zn2+活化)。可被乙二胺四羧酸和P-所抑制;第4型為不被克拉維酸抑制的抗生素酶。臨床常見的β—內丙酯酶有超低毒β—內丙酯酶、頭孢菌素酶(AmpC酶)和金屬酶。
(1)超低毒β-內丙酯酶(-β-,ESBLs)
ESBLs是一類才能酯化抗生素類、頭孢菌素類及單環類藥物的β—內丙酯酶,屬Bush分型中的2型β—內丙酯酶,其活性能被個別β—內丙酯酶抑制劑(棒酸、舒巴坦、他唑巴坦)所抑制。ESBLs主要由普通β-內丙酯酶基因(TEM—1,TEM—2和SHV—1等)突變而至,其耐藥性多由引物介導。自1983年在美國首次發覺ESBLs以來,目前已報導的TEM類ESBIs已有90多種,SHV類ESBLs少于25種。TEM型和SHV型ESBLs主要發覺于麻疹克雷伯菌和大腸埃希菌,亦發覺于變型球菌屬、普羅威登斯菌屬和其他腸球菌科真菌。
國外近些年來隨著三代阿莫西林菌素的廣泛使用,產ESBLs菌的檢出率逐年降低。NCCLs規定,凡臨床分離的大腸埃希氏菌和克雷伯氏菌均應檢測是否為產ESBLs弧菌;若形成,無論體外對第三代頭抱菌素、氨曲南的藥敏結果怎樣,均應報告對三代阿莫西林菌素及氨曲南耐藥。另外,ESBLs菌種除了對β-內丙酯類藥物有很高的耐藥率,并且對羥基香豆素類、喹喏酰基耐藥率也在60%左右,因而,臨床遇見由ESBLs造成的感染時,建議首選含β—內丙酯酶抑制劑的復方藥物劑型或酸酐培南;對于阿莫西林吡肟等四代阿莫西林,尚有爭議,按照抑菌藥的PK/PD理論,適當改變給藥劑量和給藥間隔。以使血藥含量超過真菌MIC的時間達40%給藥間隔以上,其實是有效的。
(2)阿莫西林菌素酶(AmpC酶)屆Bush分類中的1型(Ⅰ型)β—內丙酯酶。
一般將其分為由染色體介導形成的AmpCβ—內丙酯酶和由引物介導形成的AmpCβ—內丙酯酶,后者的形成菌有陰溝腸球菌、銅綠假單胞菌等,前者主要由麻疹克雷伯氏菌和大腸埃希氏菌形成。AmpC酶可作用于大多數抗生素,第一、二、三代阿莫西林菌素和單環類藥物。而第四代阿莫西林菌素、碳青霉烯類不受該酶作用。該酶不能被β—內丙酯酶抑制劑所抑制。AmpCβ—內丙酯酶的形成有2種可能:①在誘導劑存在時暫時高水平形成,當誘導劑不存在時,酶產值急劇下滑,三代阿莫西林菌素、棒酸和碳青霉烯類藥物是誘導型AmpC酶的強誘導劑;②染色體上控制酶抒發的基因發生突變,造成AmpC酶持續穩定高水平抒發。由高產AmpC酶耐藥菌造成的感染死亡率很高。
實際上,所有的革蘭氏陽性菌都能形成染色體介導的AmpC阿莫西林菌素酶,在多數情況下為低水平抒發;在腸球菌、檸檬酸鏈球菌、沙雷氏菌、銅綠假單胞菌中可高頻誘導形成,且常為高產突變株。當臨床出現上述病菌感染,開始幾天三代阿莫西林菌素醫治敏感,而此后發生耐藥時,我們可懷疑為高產AmpC酶的真菌感染,四代阿莫西林菌素和碳青霉烯類藥物不受具影響,可供臨床選用。含酶抑制劑的復方劑型不能用于醫治產AmpC酶弧菌的感染。
(3)金屬酶(β-)
大部份β-內丙酯酶的活性位點是谷氨酸殘基,但也有一小部份活性位點為金屬離子的酶類。第一個發覺的以金屬離子為活性中心的酶是由蠟樣芽抱球菌形成的阿莫西林菌素酶,能被EDTA所抑制,然后世界各地均發覺了能形成這類酶的各類病菌。1988年Bush首次將該酶定名為金屬β-內丙酯酶(β-),簡稱金屬酶。金屬β-內丙酯酶耐受β—內丙酯酶抑制劑且可酯化幾乎所有β—內丙酯類藥物(包括吡啶培南)。該酶已在氣單胞菌、嗜麥芽窄食單胞菌、洋蔥伯克霍爾德氏菌中發覺,其中嗜麥芽窄食單胞菌的吡啶培南耐藥性由染色體介導,而脆弱擬鏈球菌、肺炎克雷伯氏菌、銅綠假單胞菌中引物介導的突變株在臺灣已有報導。由粘質沙雷氏菌形成的金屬β—內丙酯酶IMP-1型可在類似接合子的intl3上聯通,早已傳播到銅綠假單胞菌、肺炎克雷伯氏菌和產堿鏈球菌。金屬酶可以酯化碳青霉烯類和近來開發的第四代阿莫西林菌素。金屬β-內丙酯酶有廣泛傳播的潛力,對幾乎所有的β—內丙酯類藥物均具有酯化活性,是目前所知的最強的β-內丙酯酶-。
2、氨基糖苷修飾酶(或鈍化酶/滅活酶)
在真菌對羥基香豆素類藥物形成耐藥的機制中,修飾酶介導的耐藥最為流行,酶促修飾的羥基香豆素類藥物不能與內質網體靶位作用,因而喪失抑菌活性。修飾酶主要包括甲基轉移酶、磷酸轉移酶和核苷轉移酶。三類甲基香豆素修飾酶的作用機制各不相同:甲基轉移酶(AAC)修飾依賴于酰基輔酶A的N-甲基化:乙酸轉移酶(APH)修飾依賴于ATP的O-乙酸化;核酸轉移酶(ANT)修飾依賴于ATP的腺苷化。在革蘭氏陽性病原菌中,最常見的甲基香豆素修飾酶是AAC(6’),使甲基香豆素類藥物1—、3—、2’—或6'—位甲基化,現在已發覺16種編碼AAC(6’)的基因。銅綠假單胞菌和腸球菌科真菌趨于于形成AAC(3)、AAC(6’)、ANT(2’’)以及APH(3’);藍莓桿菌和糞腸桿菌常常形成ANT(4’)(4’’)或雙功能的AAC(6’)/APH(2”)。獼猴桃桿菌對慶大霉素、卡那霉素和妥布霉素的`耐藥性和腸桿菌的高度慶大霉素耐藥性一般由雙功能酶介導,這種酶一般(但非總是)由坐落多重耐藥引物上的轉座子(Tn924)編碼,如藍莓桿菌具有的轉座子編碼的APH(3’)(提供卡那霉素、新霉素和阿米卡星耐藥性),而其他的定位于染色體。越來越多的菌種可形成2種或更多種酶,對抗甲基香豆素類藥物。在過去幾年里常見的組合是慶大霉素修飾酶ANT(2’’)和AAC(3)]與AAC(6’)結合,造成對慶大霉素、妥布霉素、耐替米星、卡那霉素和阿米卡星的低毒耐藥性。
甲基香豆素類藥物對非發酵菌、腸球菌科及一些革蘭氏陰性桿菌均有挺好的抑菌活性,與β—內丙酯類藥物聯用有協同抑菌作用,在感染醫治中占有重要地位。但因為以上耐藥機制的存在,真菌耐藥問題也日趨嚴重,應當導致注重,可喜的是阿米卡星等對MRSA和產ESBLs菌種仍保持17%-40%的敏感率。
二、改變抗生素作用靶位
1、青霉素結合蛋白(PBP)的改變引起的β—內丙酯類藥物耐藥
抗生素結合蛋白(PBP)參與了肽聚糖合成的最后階段。高分子量PBP經常為多模塊,具有N末端糖基轉移酶區和C末端轉肽酶區。轉肽酶區的活性位點谷氨酸與酶的天然結構相近,可與與β—內丙酯類藥物發生不可逆酰化。鏈霉素結合蛋白(PBP)的改變常造成如下兩種臨床重要的耐藥表型。
(1)耐甲氧西林金紅色獼猴桃桿菌(-,MRSA)
MRSA是20世紀60年代美國首先報導的一種嚴重的臨床耐藥致病菌,20世紀80年代以來,世界各地都陸續發生MRSA診所感染的暴發流行,并逐年增多。MRSA耐藥分為固有耐藥和獲得性耐藥,固有耐藥是由染色體介導的,其耐藥性的形成是由于真菌形成一種特殊的抗生素結合蛋白PBP2a(或PBP2’),分子量為78000的蛋白質,與β內丙酯類藥物的親和力降低,進而造成真菌對β-內丙酯類藥物耐藥。PBP2a由mecA基因編碼,95%以上的MRSA弧菌能測量到mecA基因,而敏感株則無。獲得性耐藥是由引物介導的,真菌獲得耐藥基因后,形成大量β-內丙酯酶(而不是PBPs),使耐酶抗生素平緩失活,表現出耐藥性,多為臨界耐藥。
在MRSA檢查過程中,凡屬MRSA,不管其對其他β-內丙酯類藥物MIC值或抗菌圈的大小細胞膜透性,實驗室均應向臨床報告為對所有抗生素類、頭孢菌素類、碳青霉烯類、碳阿莫西林烯類和β內丙酯類—酶抑制劑復合劑型耐藥,以免欺騙臨床服藥。MRSA感染的醫治是臨床非常棘手的困局之一,關鍵是其對許多藥物具有多重耐藥性,萬古霉素是目前臨床上醫治MRSA效果肯定的藥物,應用30多年來未發覺耐藥弧菌。
(2)耐抗生素腦炎桿菌(,PRSP)
常年以來麻疹球菌對抗生素高度敏感。MIC在0.005-0.01mg/L之間。1967年美國首次報導耐抗生素麻疹球菌,MIC為0.5mg/L,隨后世界許多國家和地區均有報導,且耐藥率迅速上升。PRSP的耐藥機制百日咳桿菌的抗生素結合蛋白(PBP)發生改變,使其與抗生素的親和力降低。百日咳桿菌有6種PBP:1a、1b、2x、2a、2b和3,其中PBP2b最為重要,假如鏈霉素結合到PBP2b上并使之抑制即造成真菌溶化和死亡;反之,PBP2b發生突變,抗生素不能形成作用,則造成PRSP。在PRSP高耐菌種中(MIC≥2μg/m1)可有多達4種PBP(主要是1a、1b、2x、2b)同時發生改變[7]。
百日咳桿菌是造成社區獲得性氣胸的重要致病菌。目前,國外PRSP的發生率在4%左右,顯著高于亞洲國家細胞膜透性,在歐洲也屬于中等水平,且MIC多大于1mg/L,為此,在社區獲得性腦部感染病原菌中,PRSP尚不構成嚴重恐嚇,抗生素仍可作為首選醫治抗生素。并且耐藥沒有國界,中國近日PRSP發生率尚低.但決不意味著不要注重,而是應當進一步強化PRSP的耐藥檢測。對于PRSP感染臨床診治推薦使用阿莫西林噻肟/阿莫西林曲松、新喹諾酸酯(如司帕沙星)。若屬PRSP嚴重感染則需應用萬古霉素或加用甲硝唑。
2、DNA拓撲異構酶的改變導致喹諾酸酯藥物耐藥
喹諾酰基抗生素的作用機制主要是通過抑制DNA拓撲異構酶而抑制DNA的合成,進而發揮抗菌和滅菌作用。真菌DNA拓撲異構酶有I、Ⅱ、Ⅲ、Ⅳ,喹諾酰基抗生素的主要作用靶位是拓撲異構酶Ⅱ和拓撲異構酶Ⅳ。拓撲異構酶Ⅱ又稱DNA促旋酶,參與DNA超螺旋的產生,拓撲異構酶Ⅳ則參與真菌子代染色質分配到子代真菌中。革蘭氏陽性菌中DNA促旋酶是喹諾酰基的第一靶位,而革蘭氏陰性菌中拓撲異構酶Ⅳ是第一靶位。
當編碼組成DNA促旋酶的A亞單位和B亞單位及組成拓撲異構酶Ⅳ的parC和parE亞單位中任一亞基的基因發生突變均可導致喹諾硫醇的耐藥性。在所有的突變型中,以gyrA的突變為主,占80%左右,其次是gyrB、parC和parE突變。在所有那些突變類型中,若Ⅱ型拓撲異構酶上存在2個突變點(如gyrA和parC上),它們造成對氟喹諾硫醇的耐藥遠遠小于只有一個突變點(如gyrA或gyrB上),后者是前者的3-4倍。同時沒有發覺突變僅出現在parC基因這一現象。這可能是由于DNA促旋酶是氟喹諾硫醇的重要靶位,gyrA亞單位的改變可導致酶結構發生變化致空間位障,制止喹諾硫醇步入喹諾酸酯作用區,或造成化學物理變化,干擾喹諾酮與酶的互相作用。這種結果顯示gyrA上突變的出現是導致真菌對喹諾酰氯發生耐藥的主要機制,而parC突變只是進一步導致銅綠假單胞菌對喹諾酮的高度耐藥。
DNA拓撲異構酶的改變是真菌耐喹諾硫醇抑菌藥的主要機制,其他耐喹諾硫醇的機制還包括前面即將提到的真菌膜私密性改變和主動外排機制。
三、細胞膜透性屏障和藥物主動外排泵
真菌可以通過細胞壁的障礙或細胞膜私密性的改變,產生一道有效屏障,致使藥物未能步入細胞內并達到作用靶位而發揮抑菌效能,這也是真菌在進化與飼養過程中產生的一種防衛機制。這類耐藥機制是非特異性的,主要見于革蘭氏陽性菌。由于革蘭氏陽性菌細胞壁粘肽層外邊存在著類脂單層組成的外膜,內層為脂寡糖,由緊密排列的碳氮分子組成,妨礙了疏水性抑菌藥步入菌體內。另外真菌外膜上還存在著多種孔蛋白,分子較大者為OmpF,分子較小者為OmpC,它們可產生特異性通道(OprD)和非特異性的通道(OprF),作為營養物質和親水性抑菌抗生素的通道。抑菌抗生素分子越大,所帶負電荷越多,疏水性越強,則不易通過真菌外膜。真菌發生突變喪失某種特異孔蛋白后即可引起真菌耐藥性,另外因為外膜蛋白OprF的缺位,使抗生素不易通過而形成耐藥性。如銅綠假單胞菌特異性孔蛋白OprD2缺位即造成碳青霉烯類藥物耐藥。
另外一種造成真菌非特異性耐藥的機制是真菌主動外排泵的存在,可以將步入真菌體內的抗生素泵出膜外,因而逃避藥物的作用。主動外排系統因為能特異地將步入細胞內的多種抑菌抗生素主動泵出細胞外,造成細胞獲得耐藥性。如大腸埃希氏菌中的多藥外排泵-TolC系統可以造成真菌對包括氯霉素、氯霉素、紅霉素、β—內丙酯類、利福平、氟喹諾硫醇、氧化劑、有機溶劑、堿性顏料等多種結構不相關的抗生素耐藥。銅綠假單胞菌的MexAB-OprM系統的主動外排作用也是造成銅綠假單胞菌固有的多重耐藥性的重要誘因之一。
真菌的膜耐藥機制主要表現在銅綠假單胞菌的多藥耐藥性。銅綠假單胞菌幾乎涵蓋了包括膜耐藥在內的所有真菌耐藥機制,其耐藥已成為當前感染醫治中較為棘手的問題之一,尤其值得注重和研究。
耐藥
互聯網