動作電位是短暫、快速的膜電位的變化(100mV),在此期間,細胞膜內(nèi)、外的極性發(fā)生反轉(zhuǎn),即細胞膜由靜息狀態(tài)時的膜內(nèi)為負、膜外為正,轉(zhuǎn)變?yōu)槟?nèi)為正而膜外為負的狀態(tài)。一個單個動作電位僅包括全部激動細胞膜的一小部份。與分級動作電位不同的是,動作電位從動作電位的起點沿整個細胞膜傳導(dǎo),傳導(dǎo)的硬度不隨距離的變化而衰減。本文介紹動作電位發(fā)生期間,細胞膜電位和離子私密性變化的一系列風(fēng)波是怎么發(fā)生的。
動作電位期間細胞膜電位的迅速反轉(zhuǎn)
假如給細胞膜一個較小的不能使其形成動作電位的電剌激,細胞膜將形成一個分級電位()。不斷降低剌激硬度,則分級電位的幅值也逐步減小,分級電位形成的是一種去極化的局部電位。
當給細胞膜一個能使其形成動作電位的閾剌激時,都會觀察到,首先出現(xiàn)一個平緩的去極化過程,當去極化達到約-55——-50mV的臨界水平時,即閾電位時,立刻形成了一個爆發(fā)的去極化過程。首先記錄到一個尖銳的向下偏轉(zhuǎn)的電位波形,達到0mV后膜電位的極性翻轉(zhuǎn)。與細胞膜外相比,此時細胞膜內(nèi)的電位為正,之后膜又迅速復(fù)極化,回到靜息電位水平。因為復(fù)極化的驅(qū)動力一般較大,致使膜電位的恢復(fù)超過了靜息電位值,形成了一個比靜息電位還負的電位(如,-80mV),即正后電位,之后才回到靜息膜電位水平(圖1)。
圖1動作電位期間膜電位的變化
從閾電位到峰值細胞膜透性,之后回到靜息水平這段迅速的電位變化稱為動作電位()。動作電位的膜極性翻轉(zhuǎn)部份(0——+30mV之間)稱為超射()。在一個給定的細胞中,動作電位的波形永遠是相同的。神經(jīng)細胞的動作電位通常僅持續(xù)1ms的時間。
細胞膜的滲透性和離子聯(lián)通的顯著變化引起動作電位的發(fā)生
細胞膜是怎樣從一個相對平衡和穩(wěn)定的靜息狀態(tài)轉(zhuǎn)弄成動作電位的?我們曉得K+是維持靜息膜電位最重要的離子,在靜息狀態(tài)時,細胞膜對K+的滲透性要遠超過Na+。但是在動作電位期間,細胞膜對K+和Na+的私密性發(fā)生了極大的變化,這種離子按其電物理梯度迅速跨膜流動,因為這種是帶電離子,因而這些流動產(chǎn)生了跨膜電壓。
動作電位期間的離子的流動主要與2種離子的通道有關(guān):即電流依賴性Na+門控通道和K+門控通道。可以將通道看作一扇門,它選擇性地開放充許離子通過,或關(guān)掉抵擋離子通過。組成通道蛋白的三維結(jié)構(gòu)的變化,決定通道是處于開放還是處于關(guān)掉狀態(tài)。已知存在3種類型的門控通道:①電壓門控通道;②化學(xué)門控通道;③機械門控通道。
1電流門控鈉離子通道和鉀離子通道
電流門控通道由帶電蛋白質(zhì)包定子成。通道周圍的電場能對處于細胞膜通道中的帶電荷部位施加使其結(jié)構(gòu)發(fā)生變化的力。通常來說,許多細胞膜蛋白相當穩(wěn)定,不受膜電流的波動而發(fā)生變化,但是通道蛋白對膜電流的變化具有高度特殊敏感性,一個極小的通道形態(tài)的改變將會喚起電位的變化,從而導(dǎo)致通道形態(tài)的轉(zhuǎn)換。
Na+通道有2種門控狀態(tài):激活態(tài)和失活態(tài)(圖2)。
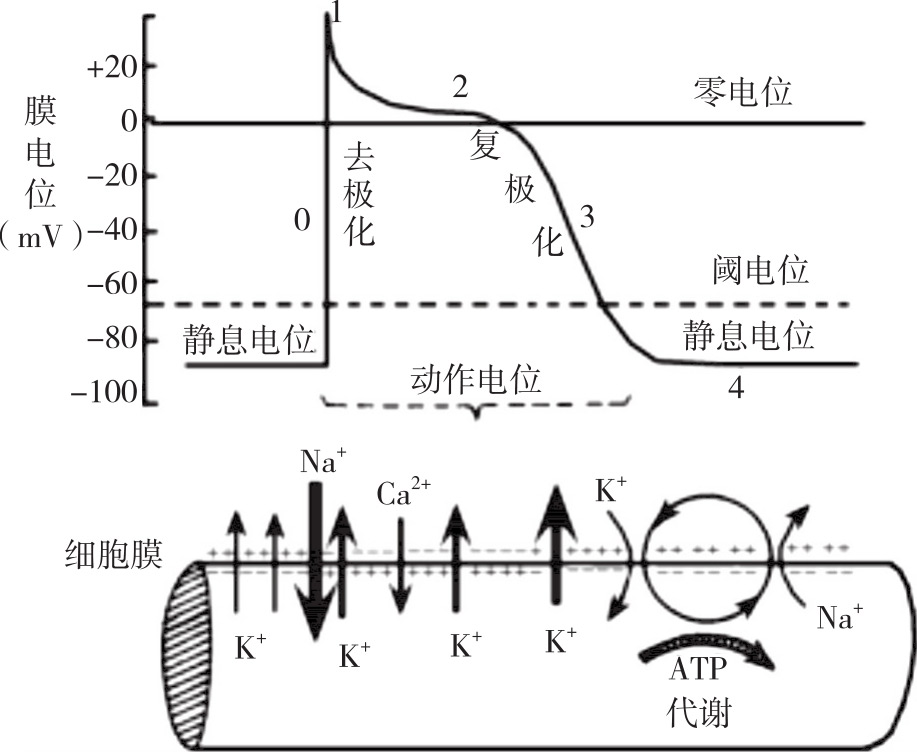
圖2電流門控Na+通道
激活態(tài)的門如同一個帶鉸鏈的門,或處于開放狀態(tài),或處于關(guān)掉狀態(tài)。失活態(tài)的門由一段多肽殘基組成,如同將一個球用鏈條聯(lián)接在一起。當球自由懸垂在鏈條的下方時,門是開放的;當球與坐落通道口的受體結(jié)合時,守門員處于關(guān)掉狀態(tài)。只有激活態(tài)和失活態(tài)2個門都處于開放狀態(tài)時,通道能夠容許離子通過。2個門中只要有任何一個處于關(guān)掉狀態(tài),離子將不能從通道通過。按此種模型,電流門控Na+通道守門員在3種狀態(tài)之間轉(zhuǎn)換:
①雖然是關(guān)掉的但有能力開放(激活態(tài)門關(guān)掉,失活態(tài)門開放);
②開放或激活狀態(tài)(2個門都處于開放狀態(tài));
③關(guān)閉態(tài),無開放的能力(激活態(tài)門開放,失活態(tài)門關(guān)掉)。
電流門控K+通道門與Na+通道門的工作原理類似,但它僅存在一種門控狀態(tài),或處于開放態(tài),或處于關(guān)掉態(tài)(圖3)。
圖3電流門控K+通道
2動作電位期間離子私密性的變化
在膜的靜息狀態(tài),所有的Na+通道和K+通道均處于關(guān)掉狀態(tài),此時,Na+通道的激活態(tài)門是關(guān)掉的,而失活態(tài)門是開放的。這表明,此時門控Na+通道雖關(guān)掉但卻處于有能力開放的狀態(tài)。在靜息狀態(tài)時沒有Na+或K+通過電流門控通道流動,但是,因為存在許多漏K+通道和極少量的漏Na+通道,靜息狀態(tài)時K+通過膜滲透能力較Na+大50——75倍,一直存在Na+和K+的透膜漏水。
隨著細胞膜向閾電位方向發(fā)展,膜去極化,一些Na+通道的激活態(tài)門開放,即此時Na+通道的2種狀態(tài)的門都處于開放狀態(tài),Na+的含量梯度(膜外低于膜內(nèi))和電流梯度(膜外為正,膜內(nèi)為負)都驅(qū)使Na+迅速向細胞內(nèi)流動,攜帶正電荷的Na+的流動使膜進一步去極化,越來越多的Na+通道開放,致使越來越多的Na+內(nèi)流,產(chǎn)生了一個正反饋過程。
去極化達到閾電位時,膜對Na+的私密性忽然顯著減小,超過了K+私密性的600倍。此時,不管是處于開放還是處于關(guān)掉狀態(tài)的通道都不再能開放。在去極化初期時相時,隨著越來越多的Na+通道的開放,膜電位開始減少,當達到閾電位時,Na+通道開放的數(shù)目早已足以啟動一個動作電位形成的正反饋進程,使余下的大量的Na+通道也陸續(xù)開放。與K+的私密性相比,此時細胞膜對Na+的私密性搶占了絕對的優(yōu)勢,大量的Na+步入細胞內(nèi),膜內(nèi)電位迅速由負變正,并接近Na+的平衡電位(約+60mV)。此時電位已達到+30mV,但并未真正達到Na+的平衡電位水平細胞膜透性,這是因為此時Na+通道開始關(guān)掉步入失活態(tài),Na+的私密性增長到靜息狀態(tài)水平。
是哪些風(fēng)波導(dǎo)致Na+通道步入關(guān)掉狀態(tài)的?當膜達到閾電位值的時侯,每位Na+通道門控變化與存在的2個緊密相關(guān)的風(fēng)波有關(guān),首先激活態(tài)門迅速開放造成膜的去極化,使通道轉(zhuǎn)換成開放的構(gòu)象(圖4)。
圖4動作電位期間離子流動和膜電位變化的關(guān)系
但是,令人不可思議的是,在通道開放的同時也啟動了通道的關(guān)掉過程,通道構(gòu)象的變化打開了通道,同時也使失活態(tài)門小球與開放態(tài)門的受體相結(jié)合,阻塞了離子通道的孔道。與迅速開放的通道相比,失活態(tài)門關(guān)掉的速率較慢。在激活態(tài)門開放以后,失活態(tài)門關(guān)掉之前的一段時間,大概存在0.5ms的延后時間,2種狀態(tài)的門都處于開放狀態(tài),Na+快速流入細胞內(nèi),造成動作電位達到峰值。以后,失活態(tài)門開始關(guān)掉,膜對Na+的私密性垂直降至靜息膜電位的水平。Na+通道維持著這些失活的構(gòu)象直到膜恢復(fù)到它的靜息值。
在動作電位達到峰值,即Na+通道失活的同時,電流門控K+通道開始開放。K+通道門對去極化的閾剌激形成的是延后性電流反應(yīng)。在閾剌激時,形成了3個相互聯(lián)系的風(fēng)波:
①Na+激活態(tài)門的迅速開放,使Na+步入到細胞內(nèi),使膜從閾電位水平迅速升至動作電位的峰值;
②Na+失活態(tài)門平緩關(guān)掉,在經(jīng)歷一個短暫的時間延后后,制止Na+的進一步內(nèi)流,使電位值不能繼續(xù)上升;
③K+通道門平緩開放,造成了動作電位從峰值返回到靜息狀態(tài)水平。
當Na+通道關(guān)掉而K+繼續(xù)從膜外向膜外滲漏時,因為沒有Na+繼續(xù)步入細胞內(nèi),膜電位必然會漸漸平緩恢復(fù)到靜息電位水平。但是,在動作電位達到峰值的時侯,因為K+通道在此時的開放,加速了膜電位向靜息電位水平恢復(fù)的速率,電流門控K+通道的開放極大地降低了K+的滲透性,是靜息狀態(tài)時Na+私密性的300倍,大量攜帶正電荷的K+從細胞內(nèi)流出,造成K+含量梯度和電位梯度在膜內(nèi)增長。值得注意的是,在動作電位峰值時,因為細胞的正電位對胞內(nèi)K+的敵視作用,此時K+的電勢梯度是從膜內(nèi)指向膜外的,與靜息電位時膜的電勢梯度的方向恰好相反。
隨著動作電位恢復(fù)到它的靜息狀態(tài),變化的膜電流使Na+通道完全關(guān)掉,此時,Na+通道的激活態(tài)門處于關(guān)掉,失活態(tài)門處于開放狀態(tài)。這是一種有能力重新開放的構(gòu)象狀態(tài),它時刻打算對到來的另一次新剌激形成反應(yīng)。動作電位期間開放的電流門控K+通道門也已關(guān)掉,只有少量漏K+通道開放,充許少量K+從細胞內(nèi)露出。因為電流門控K+通道關(guān)掉的速率較平緩,它持續(xù)降低了細胞膜對K+的滲透性,稍稍過量的K+的外流使細胞內(nèi)電位較靜息狀態(tài)時更負,產(chǎn)生一個超極化電位(圖4g)。
鉀離子通道對靜息膜電位的特殊作用:調(diào)節(jié)電激動和中止動作電位
K+通道是迄今所知分布最為廣泛的、最大的電流門控離子通道家族。腰部植物起碼有17個不同基因編碼的、具有S1——S6不同形態(tài)的K+通道。從K+通道通過的離子通常有特別高的選擇性,通透能力有較大差異,其中,K+>Rb+>NH4+》Cs+>Li+,Na+,Ca2+。在正常生理情況下,Pk/PNa(K+和Na+通透率的比值)的通透百分比小于100,并且Na+能阻斷K+通道。在完全缺K+的情況下,一些K+通道可容許Na+通過,這些特點與Ca2+通道類似。Ca2+通道在完全缺Ca2+時,也能充許Na+電壓和K+電壓通過。
按照K+通道的高度特異選擇性及平衡電位接近-90mV的特性,K+通道的最基本的作用,應(yīng)是使激動的細胞遭到抑制。K+通道對抗Na+和Ca2+通道的激動性活動,起著穩(wěn)定靜息電位的作用,使細胞保持非激動狀態(tài)。雖然一些K+通道對靜息電位起著決定性的作用,但是K+通道在可激動細胞中的電流依賴性和動力學(xué)特點,使它們還具有其他一些特殊的功能,比如調(diào)節(jié)復(fù)極化過程、修飾動作電位時程、控制沖動的領(lǐng)取頻度、決定節(jié)律性脈沖領(lǐng)取的特點等。K+通道的這種特性使其在調(diào)節(jié)所有類型胸肌收縮的硬度和頻度中,在神經(jīng)終末中止神經(jīng)遞質(zhì)的釋放中,以及在弱化突觸聯(lián)接硬度的風(fēng)波中,均發(fā)揮極為廣泛和重要的作用。
