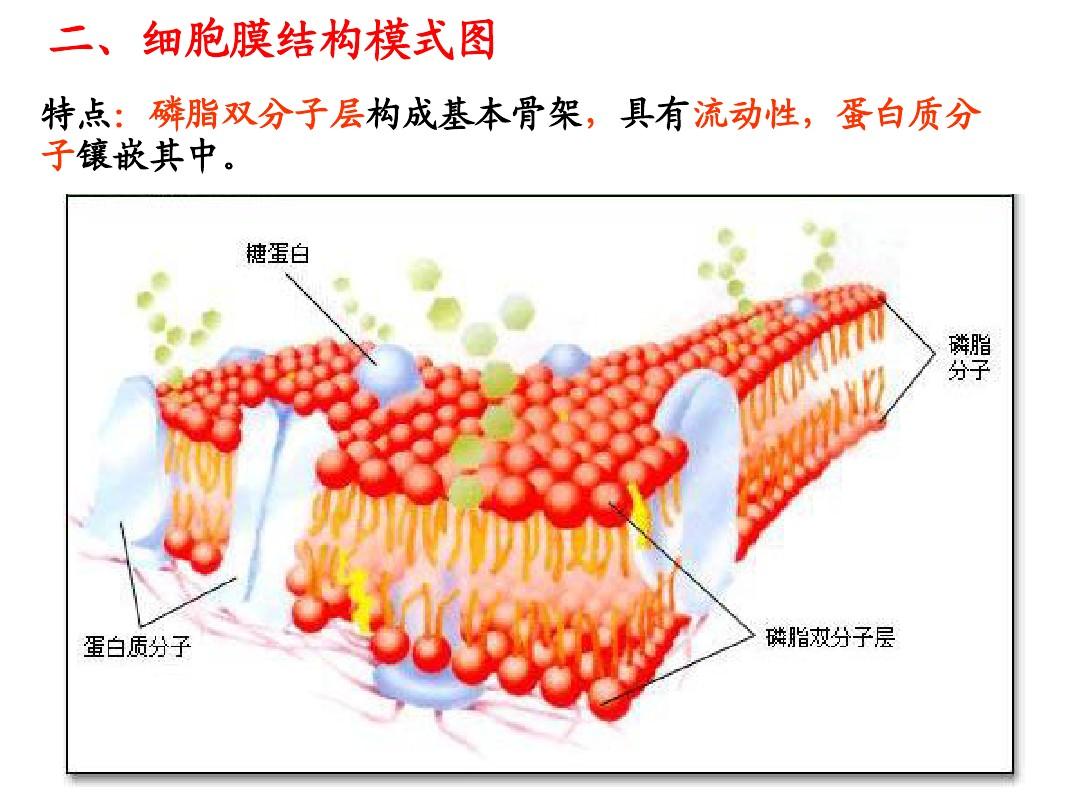
細胞膜的研究進展(—)細胞膜基礎1、細胞膜的結構任何細胞都以一層薄膜將原生質與環(huán)境分開,這層膜稱為細胞膜()或質膜()。細胞膜主要由蛋白質和脂類組成,再者還有少量的鞣質.水和金屬離子等。細胞膜的長度約7-10nm。在電鏡下觀察,細胞膜的切面圖呈暗(電子密度高)-明(電子密度低)-暗三層結構。通常覺得,電子密度高的暗層部份是由蛋白質和磷脂的腹部組成,而電子密度低的明亮區(qū)相當于磷脂的尾部(脂肪酸烴鏈)。在細胞膜結構中,兼性分子磷脂的親水性的背部都朝向膜的內、外兩表面;而疏水性的尾部都朝向膜的中央,因而產生一個連續(xù)的磷脂雙分子層,雙分子層內外兩邊各覆蓋一層蛋白質。羅伯特桑(,1959)將這些三合板式結構的膜稱為單位膜()。細胞內的各類膜系構造都與單位膜大致相同。關于細胞膜的分子結構,許多學者提出了不同的學說和模型,其中遭到廣泛支持的是和于1972年提出的“流動鑲嵌模型”()(圖1-1)圖1-1該學說覺得細胞膜是由兩層磷脂分子和嵌入的球蛋白分子構成的,磷脂雙分子層是細胞膜的主體,它不是固態(tài)物質,而是可活動的液態(tài)物質;蛋白質在磷脂雙分子層中的鑲嵌是不規(guī)則的,分布也是不對稱的,有的附在磷脂雙分子的內外表面,稱外在蛋白()或周邊蛋白(),有的則橫越膜層,稱膜內在蛋白質()。
嵌入的蛋白質分子可以進行側向擴散運動或垂直上下運動。細胞膜上的外在蛋白通常和細胞的吞噬.吞飲作用,細胞的變型運動以及細胞分裂中細胞膜的分割有關;內在蛋白則大多是轉運膜內外物質的載體、特異性的酶、受體及免疫蛋白等。在細胞膜下,具有各類免疫活性蛋白相連的,由纖維蛋白組成的網架結構,稱為膜骨架()。如喂奶植物的紅細胞骨架蛋白只要包括血影蛋白、肌動蛋白和瞄蛋白等,其他種類細胞骨架的組成還不太清楚。膜骨架參與了維持質膜的形狀以及協助質膜完成多種生理功能。2、細胞膜的功能細胞膜是細胞與外界環(huán)境的界膜,細胞與周圍環(huán)境所發(fā)生的一切聯系和反應均需依靠于膜能夠完成。為此,細胞膜具有多方面的重要功能,如物質運輸、信息傳遞、細胞辨識及免疫等。(1)物質運輸作用物質進出細胞必須通過細胞膜。細胞膜對物質的通透具有高度的選擇性,即能選擇性的透過個別物質。所以,細胞膜是一種選擇性半透膜。細胞膜的這些特點對維持細胞的正常生理功能是極其重要的。物質分子通常總是從高含量區(qū)間向低含量區(qū)間方向擴散,最后趨向平衡,這些現象成為擴散作用()。分子通過一層像細胞膜這樣的半透膜進行擴散,稱作滲透作用()細胞膜是一種選擇性半透膜,細胞膜對物質的通透是按細胞膜內外各類物質的特點及含量進行的有選擇地、可變地通透,物質分子由高含量處經細胞膜向低含量處的通透稱為被動運輸()。
被動運輸包括自由擴散和推動擴散,如一些脂胺類物質、水、糖類、氨基酸等的通透。而又、由低含量處經細胞膜向高含量處的運輸,則稱為主動運輸()。主動運輸是一個耗能量并須要載體(即膜上的嵌入蛋白分子)協助的運輸過程。如人體細胞通過細胞膜上的嵌入蛋白——Na泵逆著濃度梯度在細胞內不斷積累離子而排出Na離子的主動運輸。主動運輸可使細胞內一些多肽的濃度高出細胞外近10倍或更多。細胞膜對于大分子物質是不能滲透的。這種物質是以產生水泡的形式進出細胞,稱內吞()或外排作用(),內吞的物質為固態(tài)者稱為吞噬(),內吞的物質為液體者稱為胞飲()。內吞作用和外排作用與其他主動運輸一樣也須要能量供應。(2)信息傳遞作用生物體內的激素、神經遞質和抗生素對細胞有調節(jié)作用,都與細胞膜有密切關系。通常覺得她們是與細胞膜上各類特異性受體()(膜蛋白)結合,通過激活受體而使信息進行跨膜轉移,最后正常抵達細胞內部而形成代謝反應。(二)細胞膜的發(fā)覺1855年,耐格里發(fā)覺色素透入已損傷和未損傷的動物細胞的情況并不相同。
他便通過細胞的滲透特點去研究它的“邊界”(他首次把細胞“邊界”稱為“質膜”)。耐格里和克拉默()一起進行實驗,通過實驗發(fā)覺細胞具有敏感的滲透特點,它的容積可以隨著周圍介質的不同滲透硬度而改變。當細胞外邊的溶質滲透硬度大時,細胞就變小;溶質滲透硬度小時,細胞就變大。耐格里提出,細胞與環(huán)境之間正是通過這些“邊界”發(fā)生關系的。耐格里在試驗中還發(fā)覺這樣的情況:把麗藻屬()長導管細胞的一端裝入水堿液內,另一端放進糖堿液,細胞內含物發(fā)生了傳動障礙。在水底一端的細胞樹汁流向糖氨水中的一端,并帶著所有可聯通的粒子。而且細胞膜蛋白,原來已知的事實表明,蒸騰作用和滲透壓加在一起也不足以將液體壓到動物的下部,這兩種力難以解釋動物樹汁流動的方向。因此耐格里覺得,不得不假定有一股其他的力量,它們在縱壁,更可能在橫壁上。這些力量加強了細胞堿液從下往上的流向。據悉,美國動物生理學家普費弗(W.)對動物細胞的滲透行為進行了大量的試驗,并于1897年提出了兩個重要的推論:第一,細胞是被質膜包被著的;第二,這層質膜是水和溶質通過的普遍障礙。同時,很快又發(fā)覺,細胞膜這個屏障具有顯著的選擇性,一些物質可通過它,而另一些物質幾乎完全不能通過。
1899年,西班牙細胞生理學家奧弗頓(C.)發(fā)表一系列關于化合物步入細胞的觀察結果,他發(fā)覺分子的極性越大,步入細胞的速率越小,當降低非極性官能團(如酰基鏈)時,化合物步入的速率便降低。奧弗頓的推論是,控制物質步入細胞的速率的細胞膜是脂肪性物質,其中富含脂類和其他醇類。為此,當時確立了有一層脂類的膜圍繞著細胞的認識。到1925(F.)又提出脂類膜具有雙分子層的概念。雖然,學者們對膜的狀況的認識都還是假定,她們都無法觀察到細胞膜。其實這個時期組織標本的固定和染色方式有了進展,甚至出現相差顯微鏡和干涉顯微鏡,但仍區(qū)分不出細胞膜來。雖然最好的光學顯微鏡也難以達到這個目的。1930—1950年,隨著電子顯微鏡技術的發(fā)展,當應用這項技術來研究細胞時,才發(fā)覺細胞的邊界膜是一個固體結構的實體,因而否認了細胞膜的存在。電鏡觀察表明,細胞遠不是一個具有核和一些懸浮在原生質膠凍中的線粒體口袋,而是一個有膜包被著的許多膜的集聚體。50年代早期,帕拉德(G.E.)和波特(K.R.)稱這些廣泛的細胞內膜系統為葉綠體。初期的電鏡工作所者觀察到的細胞內的各類膜與“有軌電車軌道”和“鐵路軌道”的隱喻大體相像。
按照細胞的生理生化特點,曾先后推論質膜是一種脂肪柵、脂類雙分子層和由蛋白質-磷脂-蛋白質構成的三夾板結構。同時電鏡觀察也否認質膜確實呈暗-明-暗三層結構。隨即冷藏刻蝕技術顯示單層膜中存在蛋白質顆粒;免疫螢光技術證明質膜中蛋白質是流動的。據此S.J.等人在1972年提出生物膜的流動鑲嵌模型,如圖1-2,結構特點是:生物膜的骨架是磷脂質雙分子層,蛋白質分子以不同的形式鑲嵌其中,細胞膜的表面還有脂類分子,產生糖脂、糖蛋白;生物膜的內外表面上細胞膜蛋白,糖類和蛋白質的分布不平衡,反映了膜外側的功能不同;脂單層具有流動性,其鹽類分子可以自由聯通,蛋白質分子也可以在脂單層中縱向聯通。雖然目前還沒有一種才能直接觀察膜的分子結構的較為便捷的技術和技巧,但從研究中30年代以來提出了各類假說有數十種,其中得到較多實驗事實支持而目前仍為大多數人圖1-2所接受的是德國的S.J.1972年提出的流體鑲嵌模型(fluidmodel)。這一假想模型的基本內容是:膜的共同結構特征是以液態(tài)脂類雙分子層為基架,其中鑲嵌著具有不同分子結構、因而也具有不同生理功能的蛋白質,前者主要以α-螺旋或球型蛋白質的方式存在。
其局限性在于未抒發(fā)出流動性不均一,Jain與White提出了“板塊與鑲嵌模型”。1、單位膜模型(1)概述單位膜模型,又叫unitmodel。1959年J.D.所提出。主要是根據電子顯微鏡的觀察,發(fā)覺細胞膜是類似鐵軌結構(“track”),兩條明線被一條明亮的帶隔開,顯示暗---明---暗的三層,總長度為7.5nm,中間層為3.5nm,內外兩層各為2nm。并猜想:暗層是蛋白質,透明層是脂,并建議將這些結構稱為單位膜。(2)特征主要是依據電子顯微鏡的觀察,發(fā)覺細胞膜是類似鐵軌結構(“track”),兩條明線被一條明亮的帶隔開,顯示暗---明---暗的三層,總厚度為7.5nm,中間層為3.5nm,內外兩層各為2nm。并推論:暗層是蛋白質,透明層是脂,并建議將這些結構稱為單位膜。單位膜模型是在片層結構模型的基礎上發(fā)展上去的另一個重要模型。它與片層結構模型有許多相同之處,最重要的更改是膜脂雙分子層內外兩邊蛋白質存在的方式不同。單位膜模型指出的是蛋白質為雙層伸展的β折疊條狀,而不是球狀蛋白。
另外,單位膜模型還覺得膜的內側表面的膜蛋白是糖蛋白,但是膜蛋白在兩側的分布是不對稱的。這一模型就能解釋細胞質膜的一些基本特點,比如質膜有很高的阻值,這是因為膜脂的非極性端的碳溴化合物是不良導體的緣故;再如因為膜脂的存在,使它對脂胺類強的非極性分子有較高的私密性,而脂胺類弱的小分子則不易透過膜。(3)發(fā)展情況隨著電鏡的問世和技術的不斷改良,用電鏡直接觀察到了膜的結構:細胞膜呈暗一明一暗的三層結構,膜長度為75A。覺得:電鏡下的明區(qū)相當于脂分子的非極性尾部,而暗區(qū)則相當于蛋白質和脂分子的極性腹部區(qū)域,蛋白質坐落脂雙分子層內外表面、為雙層伸展的β一片層。并提出所有細胞膜在電鏡下都具有這些暗—明—暗三層結構,且膜長度為75A左右,稱之為單位膜。單位膜模型是在片層結構模型的基礎上發(fā)展上去的另一個重要模型。它與片層結構模許多相同之處,最重要的更改是膜脂雙分子層內外兩邊蛋白質存在的形式不同。單位膜模型指出的是蛋白質為雙層伸展的β折疊條狀,而不是球狀蛋白。另外,單位膜模型還覺得膜的內側表面的膜蛋白是糖蛋白,并且膜蛋白在兩側的分布是不對稱的。
這一模型還能解釋細胞質膜的一些基本特點,比如質膜有很高的內阻,這是因為膜脂的非極性端的醇類化合物是不良導體的緣故;再如因為膜脂的存在,使它對脂胺類強的非極性分子有較高的私密性,而脂胺類弱的小分子則不易透過膜。單位膜也有一些不足首先該模型把膜看成是靜止的,未能說明膜怎樣適應細胞生命活動的變化;其一,不同的膜其長度不都是7.5nm,通常在5~10nm之間;其一,假如蛋白質是伸展的,則不能解釋酶的活性同構型的關系。還有,該模型也不能解釋為何有的膜蛋白很容易被分離,有些則很難。不足之處:1)把膜結構描寫成靜止不變的;2)未能解釋膜的功能活動;3)各類膜有各自的特定長度,并不都是7.5nm;4)蛋白質提取的難易程度不同;5)各類膜的蛋白質和脂質的百分比不同。參考文獻2、流動鑲嵌模型(1)概述1972)和尼克森(G.)總結了當時有關膜結構模型及各類研究新技術的成就,提出了生物膜的流動鑲嵌模型(fluidmodel)。流動鑲嵌模型(fluidmodel),特指膜的流動鑲嵌模型。這一模型是針對細